Daniel J. Nicholson, noviembre de 2013
Parte I
Las partes y el todo
Esto nos lleva a otra diferencia fundamental entre los organismos y las máquinas, que se refiere a la naturaleza de la relación entre las partes y el todo. En una máquina, las partes son causalmente independientes del todo que constituyen y le son temporalmente anteriores. Sus funciones pertenecen al diseño original de la máquina y, por lo tanto, son asignadas de antemano por el diseñador de la máquina. Aunque las partes adquieren su función en virtud de estar presentes en la máquina como un todo, conservan sus propias propiedades distintivas independientemente de si están integradas en la máquina o no. Por el contrario, las partes de un organismo no son ni causalmente independientes del todo que constituyen, ni le son temporalmente anteriores. Existen en una relación de interdependencia colectiva, ya que cada parte es necesaria para la generación y el funcionamiento de las demás. El organismo mantiene su autonomía como todo regulando, reparando y regenerando constantemente sus partes. Los procesos causales por los que se producen las partes son precisamente aquellos por los que constituyen y mantienen la organización del todo. Como resultado, la generación, las propiedades y las funciones de las partes de un organismo, a diferencia de las de una máquina, no pueden entenderse independientemente del todo. Aunque el argumento no puede desarrollarse aquí, esta es una de las principales razones por las que el reduccionismo explicativo tiene una eficacia limitada a la hora de explicar los fenómenos de los organismos.
La influencia determinante del todo sobre las partes en los organismos ha sido reconocida desde hace tiempo por los biólogos. Uno de los estudios más influyentes sobre las capacidades «holísticas» de los organismos fue realizado por Goldstein (1995 [1934]), quien, en el curso de su tratamiento de soldados con daño cerebral durante la Primera Guerra Mundial, observó que el organismo se reajustaba para hacer frente a lesiones devastadoras retirándose a rangos de actividad más limitados que podía gestionar mediante la redistribución adecuada de sus energías reducidas. Para Goldstein, la suposición de que en un organismo, al igual que en una máquina, las partes determinan el todo podía refutarse sobre bases empíricas. Al enfrentarse a una enfermedad o lesión, Goldstein descubrió que los organismos poseen la flexibilidad inherente para reorganizar sus partes en gran medida con el fin de recuperar el desempeño de las funciones vitales. De este modo, el rechazo de Goldstein al OCM [Organismo Como Máquina] no estuvo motivado por consideraciones filosóficas, sino por observaciones clínicas. Incluso alguien con sólidas credenciales mecanicistas como von Neumann (quien utilizó el OCM como base para su teoría de los autómatas autorreproductores) reconoció la influencia determinante del todo sobre las partes en sistemas biológicos como el cerebro, señalando que:
«Nunca es muy sencillo localizar nada en el cerebro, porque el cerebro tiene una enorme capacidad de reorganizarse. Incluso cuando se ha localizado una función en una parte concreta del mismo, si se extirpa esa parte, es posible que se descubra que el cerebro se ha reorganizado, ha reasignado sus responsabilidades y la función se está desempeñando de nuevo» (von Neumann, 1966, p. 49).
La determinación de las partes por parte del organismo en su conjunto se extiende no solo a su función y comportamiento, como muestran los ejemplos anteriores, sino también a sus propiedades estructurales. En palabras de Haldane (1884, p. 37):
«lo que parecía pertenecer a las partes independientemente de su relación con el todo, por ejemplo, su tamaño, forma y estructura, no es en realidad más que la manifestación en las partes de la influencia del todo».
Frankhauser ofreció un magnífico ejemplo de ello en la década de 1940, tal y como nos han recordado recientemente Kirschner et al. (2000). Frankhauser llevó a cabo experimentos sobre los efectos de la ploidía (es decir, el número de juegos cromosómicos en la célula) en el desarrollo de las salamandras, y descubrió que los embriones poliploides, generados al suprimir las divisiones celulares tempranas, tenían menos células, pero estas eran más grandes. El número y el tamaño de las células diferían en los embriones haploides, diploides y pentaploides, pero los tejidos del organismo, así como el organismo en su conjunto, mantuvieron el mismo tamaño en todos los casos. Estos ejemplos de desarrollo regulativo se observaron con mayor claridad en estructuras bien definidas como el conducto pronéfrico del riñón. Los experimentos de Frankhauser demostraron que, en un organismo, a diferencia de lo que ocurre en una máquina, no existe una correlación estricta entre el número y el tamaño de las partes (en este caso, las células) y el tamaño del conjunto. Mientras que en una máquina el tamaño, la forma y la estructura del todo vienen determinados invariablemente por los de sus partes, en un organismo el tamaño, la forma y la estructura de las partes no bastan para explicar los del todo, dado que el todo ejerce una influencia determinante sobre sus partes.
Metabolismo
También es importante destacar que un organismo, a diferencia de una máquina, presenta una identidad estructural transitoria. Los materiales que lo componen cambian, pero la organización del todo se mantiene. Mientras que una máquina siempre está compuesta por los mismos componentes materiales (a menos que interfiera un agente externo), un organismo se mantiene naturalmente en un estado de flujo continuo en el que se produce una descomposición y sustitución permanentes de sus materiales constituyentes. Este es el proceso característico de los sistemas vivos que llamamos metabolismo. No tiene sentido identificar un organismo a lo largo del tiempo con la suma de sus partes materiales, ya que estas son repuestas constantemente por el todo. Las partes de un organismo en un momento dado son solo la manifestación temporal de la unidad organizativa autoproducida del todo. Las partes de una máquina, sin embargo, permanecen distintas, estables e identificables a lo largo del tiempo.
Desarrollo
Otra diferencia entre los organismos y las máquinas se refiere a la diferente prioridad ontogénica de las partes y el todo. En una máquina, el todo solo llega a existir después de que todas las partes hayan sido ensambladas adecuadamente por su fabricante. En un organismo, la existencia de las partes no precede a la del todo, dado que las partes solo adquieren sus respectivas identidades en cuanto partes a medida que el todo se desarrolla progresivamente a partir de un sistema originalmente indiferenciado, pero ya integrado. Esto apunta a otra diferencia más entre los organismos y las máquinas, que es que las actividades fisiológicas de un organismo ya deben estar teniendo lugar mientras el crecimiento está en curso, mientras que una máquina no puede realizar las funciones para las que fue diseñada mientras aún se encuentra en proceso de construcción. El organismo mantiene incluso su autonomía organizativa cuando produce descendencia o cuando se divide por reproducción asexual. Es difícil concebir una máquina que pudiera dividirse un número indefinido de veces y, sin embargo, ser capaz de conservar su unidad y funcionalidad de la misma manera que lo hacen incluso los microorganismos más simples.
Servomecanismos
Hasta ahora he defendido que la diferencia más general entre los organismos y las máquinas es que los primeros tienen una finalidad intrínseca, mientras que las segundas tienen una finalidad extrínseca. Todas las diferencias específicas entre organismos y máquinas que he analizado posteriormente están relacionadas, de una forma u otra, con esta distinción clave. Como se ha explicado anteriormente, el tipo de finalidad que muestra un sistema viene determinado por su dinámica organizativa interna. La intencionalidad intrínseca de los organismos se basa en el hecho de que están autoorganizados, se autoproducen, se automantienen y se autorregeneran. Por el contrario, la intencionalidad extrínseca de las máquinas se basa en el hecho de que están organizadas, ensambladas, mantenidas y reparadas por agentes externos.
Ahora bien, es cierto que los servomecanismos (es decir, las máquinas controladas por retroalimentación negativa) muestran patrones de comportamiento que se asemejan a los que exhiben los sistemas intrínsecamente intencionales, como la autorregulación (p. ej., un calefactor con termostato) y la autogestión (p. ej., un torpedo que busca un objetivo). Esto ha llevado a algunos autores (McShea, 2012; Nagel, 1979; Rosenblueth, Wiener y Bigelow, 1943) a sostener que los servomecanismos son indistinguibles de los organismos desde el punto de vista de su intencionalidad. Sin embargo, este argumento equipara erróneamente la intencionalidad con el comportamiento. Aunque el patrón de comportamiento de un sistema ofrece una buena evidencia de su intencionalidad, la intencionalidad de un sistema no puede explicarse en términos de comportamiento observable —y mucho menos definirse en términos de entradas y salidas mientras se trata como una caja negra la organización interna del sistema causalmente responsable de ella. Sin embargo, esto es precisamente lo que hicieron los primeros cibernéticos cuando propusieron este argumento (véase Rosenblueth y Wiener, 1950; Rosenblueth et al., 1943), al afirmar que si un perro servomecánico se comporta como un perro vivo, entonces debe atribuirse el mismo tipo de intencionalidad tanto al organismo como a la máquina «si queremos ser coherentes» (Rosenblueth y Wiener, 1950, p. 323). De ello concluyeron que «como objetos de investigación científica, los seres humanos no difieren de las máquinas» (Rosenblueth y Wiener, 1950, p. 326). El problema de esta afirmación es que la capacidad de simular fielmente un patrón particular de comportamiento orgánico en una máquina no proporciona una base legítima para concluir que la máquina es análoga al organismo que imita, dado que el mismo resultado conductual puede producirse por medios radicalmente diferentes [8].
La intencionalidad de un sistema no depende de sus patrones de respuesta conductual, sino del régimen organizativo interno causalmente responsable de ellos. Los servomecanismos carecen de la organización autosuficiente que les permita actuar genuinamente por sí mismos. Esto queda ejemplificado por el hecho de que los termostatos y los torpedos solo simulan un comportamiento intrínsecamente intencional (y solo en un sentido muy restringido) cuando están encendidos; dejan de hacerlo cuando se apagan. Sin embargo, independientemente de si están encendidos o apagados, existen (como cualquier otra máquina). Esto se debe a su organización interna. Comparemos esto con la situación de los organismos. Los organismos no pueden «apagarse» sin perder su integridad estructural. Una vez más, esto se debe a su organización interna. Una de las características distintivas de la organización autónoma de un organismo es que su funcionamiento ininterrumpido es una condición necesaria para su existencia continuada. Esta necesidad de mantener constantemente la compleja red de procesos causales responsables de la integridad estructural de un organismo es lo que subyace a su intencionalidad intrínseca. Los servomecanismos pueden simular (con diversos grados de éxito) ciertos comportamientos de los sistemas intrínsecamente intencionales, pero no son en sí mismos sujetos de acción intencional porque su actividad no está dirigida a su propia preservación. En cambio, su organización y funcionamiento reflejan los fines específicos de sus creadores y usuarios (cf. Jonas, 2001; Oparin, 1961; Taylor, 1950) [9].
Los organismos son sistemas autónomos. Las máquinas (incluidos los servomecanismos) no son más que medios para potenciar la autonomía causal de sus usuarios. Los organismos carecen de controles externos, mientras que «una característica de todas las máquinas creadas por el hombre es que sirven como instrumentos de control» (Kapp, 1954, p. 93). Las máquinas se controlan para que funcionen de la manera deseada por sus creadores y usuarios. Somos nosotros, como creadores y usuarios de las máquinas, quienes determinamos las normas de su funcionamiento. Cuando las máquinas no funcionan como esperamos, las consideramos (nosotros) defectuosas o con un mal funcionamiento. Los organismos, por el contrario, funcionan según sus propias normas. El funcionamiento de un organismo es intrínsecamente relevante para sí mismo, dado que su propia existencia depende de que su funcionamiento sea tal y como es. Un organismo (así como sus partes) debe funcionar de acuerdo con las normas operativas particulares que le permiten mantener su organización autoproducida a lo largo del tiempo; si deja de seguir estas normas (que son diferentes para cada organismo), deja de existir (cf. Barham, 2012; Canguilhem, 1991; Saborido et al., 2011).
Por último, cabría mencionar el hecho de que los organismos se producen de forma natural, mientras que las máquinas se crean artificialmente. Sin embargo, no considero que esta sea una diferencia tan importante porque, a diferencia de todas las demás discutidas en esta sección, no depende necesariamente de la distinción entre intencionalidad intrínseca y extrínseca. Aunque es cierto que hasta ahora no se ha creado artificialmente ningún organismo [10], si los biólogos sintéticos lograran finalmente diseñar desde cero un sistema vivo —es decir, un sistema que fuera autoorganizado, autoproducido (desde su creación inicial), se mantuviera a sí mismo y se regenerara por sí mismo—, entonces dicho sistema, a pesar de su origen artificial, seguiría teniendo la capacidad, en virtud de su dinámica organizativa interna, de actuar por cuenta propia de acuerdo con sus propias normas. Sostengo que la distinción entre formas intrínsecas y extrínsecas de intencionalidad es mucho más adecuada que la distinción entre orígenes naturales y artificiales para captar las características respectivas de los organismos y las máquinas.
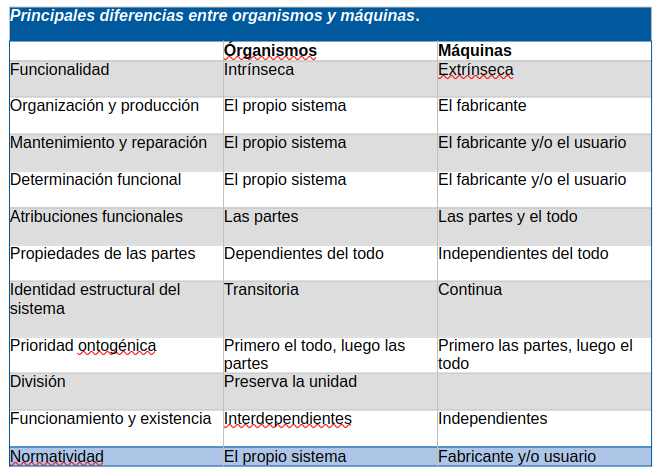
Volviendo al OCM, ahora podemos ver que su insuficiencia se deriva del problema de proyectar las características de los sistemas extrínsecamente intencionales sobre los intrínsecamente intencionales. La Tabla 1 muestra una selección de las diferencias clave entre organismos y máquinas discutidas en esta sección, que ponen de manifiesto las diversas deficiencias del OCM.
4. ¿Qué papel debería desempeñar el modelo del organismo como máquina en la biología actual?
Dadas las numerosas deficiencias del modelo del organismo como máquina, cabe preguntarse: ¿por qué ha ejercido esta noción una influencia tan poderosa en la biología? Se pueden identificar varias razones. En primer lugar, dado que las máquinas son familiares y se comprenden bien, al ser productos del diseño humano, constituyen modelos intuitivamente convincentes para conceptualizar los organismos. Como resultado, los biólogos suelen verse tentados a recurrir a las similitudes superficiales entre máquinas y organismos (por ejemplo, organización jerárquica, partes funcionales, comportamiento intencional) para explicar a estos últimos basándose en su familiaridad con las primeras. En segundo lugar, al defender el OCM, los biólogos han mantenido su disciplina firmemente dentro de los límites de la ciencia física, y esto ha servido históricamente para garantizar la respetabilidad científica de sus investigaciones, así como para legitimar la transferencia epistémica de teorías, conceptos y métodos desde ciencias más desarrolladas y «duras», como la física y la química, así como la ingeniería [11]. En tercer lugar, el OCM llena el vacío causado por la ausencia de una definición de vida generalmente aceptada, de modo que, al respaldarlo, los biólogos no han tenido que preocuparse por cuestiones espinosas como «¿Cuál es la naturaleza de la vida?» o «¿Qué es un organismo?», y en su lugar han podido dedicarse a estudiar los sistemas vivos. Pero, sobre todo, la razón más importante por la que el OCM ha sido, y sigue siendo, una noción tan seductora para los biólogos es que tiene un gran éxito a la hora de generar datos empíricos. Es innegable que los períodos de la historia en los que la tradición de investigación mecanicista ha dominado la investigación biológica han sido también los períodos de mayor progreso empírico. Esto nos lleva a una paradoja intrigante: ¿cómo puede el OCM dar lugar a una comprensión tan profundamente problemática de lo que son los organismos y, sin embargo, resultar tan fructífero cuando se utiliza para investigarlos? A la luz de esta tensión, ¿qué papel debería desempeñar el OCM en la biología actual?
Para abordar estas cuestiones, es necesario dar un paso atrás y considerar en términos generales las diferentes formas en que se utilizan las metáforas en la ciencia. Siguiendo a Bradie (1999), podemos distinguir funciones teóricas, heurísticas y retóricas para las metáforas en la ciencia. Las metáforas con una función teórica son fundamentales para la comprensión científica, ya que proporcionan la base para la conceptualización, representación y explicación del fenómeno en cuestión. Las metáforas con función heurística constituyen herramientas metodológicas que facilitan la investigación empírica del fenómeno en cuestión. Y las metáforas con función retórica se emplean en la comunicación científica para informar y educar a los no especialistas sobre el fenómeno en cuestión. Las metáforas científicas más extendidas, como el OCM, desempeñan las tres funciones. Hasta ahora, en este artículo, mi análisis del OCM se ha centrado en su función teórica, y espero que lo dicho en la sección anterior sea suficiente para concluir legítimamente que, como teoría del organismo, el OCM no tiene ningún papel que desempeñar en la biología. A continuación, examinaré sucesivamente las funciones heurística y retórica del OCM. En primer lugar, explicaré por qué el OCM tiene éxito cuando se emplea de forma heurística pero no teórica, y a continuación ilustraré los graves problemas que surgen del recurso retórico al OCM.
La función heurística
La clave del valor heurístico del OCM reside en el hecho de que la naturaleza del organismo solo es perceptible cuando se considera como un todo. Si las partes de un organismo se consideran independientemente del todo a efectos de su investigación, se asemejan a las máquinas en que constituyen sistemas extrínsecamente finalizados. Al igual que las máquinas, las partes de un organismo no se autoorganizan, autoproducen, automantienen ni autorreparan, sino que dependen de un agente externo para su organización, producción, mantenimiento y reparación —a saber, el organismo en su conjunto—. Al igual que las máquinas sirven a los fines de sus usuarios, las partes de un organismo sirven a los fines del todo al que pertenecen, razón por la cual se pueden atribuir funciones a ambos. Así pues, mientras que un organismo en su conjunto es un tipo de sistema fundamentalmente diferente de una máquina, sus partes comparten en realidad muchos de los atributos de las máquinas. En consecuencia, cuando estas partes se estudian independientemente del todo, se puede aprender mucho sobre ellas tratándolas como si fueran máquinas. Esto, creo, es la fuente del poder heurístico del OCM, y explica por qué esta noción ha demostrado ser tan exitosa cuando se utiliza como herramienta metodológica en la caracterización analítica de los organismos. Investigar áreas localizadas dentro del organismo como si fueran máquinas permite a los biólogos abstraerse convenientemente de la intimidante complejidad del contexto fisiológico más amplio del organismo en su conjunto, y centrar su atención en partes interactuantes bien definidas (cf. Bechtel y Richardson, 1993; Kauffman, 1970; Wimsatt, 1976).
No obstante, es necesario señalar una limitación importante del uso metodológico del OCM. Se trata de que el valor heurístico del OCM es directamente proporcional al grado de diferenciación fisiológica del organismo objeto de estudio. Un organismo en las etapas iniciales de desarrollo no puede estudiarse como un conjunto de subunidades mecánicas porque en él todas las funciones siguen siendo asumidas por el organismo en su conjunto. El organismo en este punto constituye lo que Driesch denominó un «sistema armonioso-equipotencial» que es prácticamente ininteligible en términos mecánicos [12]. Solo con la diferenciación progresiva del embrión la acción originalmente unitaria del organismo se divide en una miríada de acciones individuales, y es en esta etapa posterior del desarrollo cuando comienzan a surgir estructuras locales de tipo máquina dentro del organismo, lo que confiere al OCM un valor heurístico cada vez mayor. Aun así, es importante señalar que, incluso en la etapa adulta, los organismos conservan un cierto grado de plasticidad y una capacidad significativa para reorganizar sus partes y reasignar sus necesidades funcionales en consecuencia, a fin de compensar las perturbaciones externas, tal y como ilustran las investigaciones clínicas de Goldstein comentadas en la sección anterior. Por lo tanto, en ninguna etapa del ciclo de vida de un organismo es absoluta la utilidad heurística del OCM.
Hasta ahora he defendido que el valor heurístico del OCM reside en las similitudes relativas entre las máquinas y las partes de los organismos. Pero, ¿qué debemos pensar de la aplicación del OCM al estudio de los organismos completos? Los organismos completos, a diferencia de sus partes, constituyen sistemas intrínsecamente teleológicos. Por consiguiente, no pueden ser mecanizados con fines de investigación de la misma manera que sus partes cuando se consideran de forma aislada. El enfoque mecánico es intrínsecamente incapaz de abordar las propiedades sistémicas que hacen que los organismos completos sean distintivos en primer lugar. Lo único que puede hacer es centrarse en regiones localizadas y altamente diferenciadas dentro de los organismos de forma individual. ¿Significa esto que el OCM es inútil en la investigación de organismos completos? No del todo. El OCM aún puede desempeñar un papel contrafactual útil desde el punto de vista heurístico, al ayudar a resaltar lo que los organismos no son. En este sentido, el OCM sirve como un modelo falso que, en virtud de su insuficiencia inherente, puede orientar a los biólogos hacia la naturaleza real de los organismos (véase Wimsatt, 1987). Como observó Rosen (1991, p. 248):
«En definitiva, la metáfora cartesiana del organismo como máquina ha demostrado ser una buena idea. Las ideas no tienen por qué ser correctas para ser buenas; solo es necesario que, si fallan, lo hagan de una manera interesante».
En general, la clave para evaluar eficazmente el OCM es delimitar claramente su función heurística de su función teórica. Las investigaciones mecánicas exitosas de los organismos no permiten explicaciones mecánicas exitosas de los mismos. Así pues, aunque la OCM proporciona un medio conveniente para simplificar pragmáticamente la realidad biológica con el fin de facilitar su investigación, no logra, sin embargo, proporcionar una comprensión teórica adecuada de esa realidad. Sostengo que la lamentable prevalencia de la OCM en la teoría biológica contemporánea es, en gran medida, el resultado de inferir injustificadamente su veracidad ontológica basándose en su utilidad metodológica.
La función retórica
Pasemos ahora a la función retórica del OCM. Como se ha indicado anteriormente, las metáforas con función retórica se utilizan con el fin de comunicar conocimientos científicos, tanto técnicos como divulgativos, a personas no especializadas. Algunos biólogos han argumentado que el OCM, a pesar de ser teóricamente inadecuado, puede seguir teniendo un gran valor retórico porque proporciona una ayuda visual muy cautivadora a la hora de enseñar a personas no especializadas las características y propiedades de los organismos. Por ejemplo, Konopka (2002, p. 399) señala que, aunque «la metáfora de la máquina está abocada al fracaso como herramienta científica seria […] puede seguir siendo una herramienta pedagógica extraordinariamente útil en la educación de no especialistas dotados en centros juveniles locales (incluidas nuestras universidades) de todo el mundo». Esta visión, sin embargo, es problemática. Del mismo modo que resulta tentador para los biólogos inferir erróneamente la adecuación teórica del OCM basándose en su valor heurístico, el uso retórico del OCM por parte de los biólogos puede llevar inadvertidamente a los no especialistas a asumir que los organismos son realmente máquinas. En este sentido, las metáforas retóricas son armas de doble filo; del mismo modo que pueden mejorar la comprensión científica, también pueden servir para obstaculizarla (Quale, 2002). Esto depende claramente de la elección de las metáforas utilizadas; las metáforas teóricamente engañosas como el OCM hacen mucho más daño que bien a este respecto. Esto puede ilustrarse considerando las repercusiones del reciente recurso retórico a las «máquinas moleculares» por parte de los biólogos moleculares.
En 1998, Alberts, entonces presidente de la Academia Nacional de Ciencias, editó un número especial de la revista Cell (volumen 92, número 3) que reunía una serie de artículos en los que se abogaba sistemáticamente por la adopción del lenguaje de máquinas en la descripción de grandes complejos proteicos. En su introducción al número, titulada «La célula como conjunto de máquinas proteicas: Preparando a la próxima generación de biólogos moleculares», Alberts señaló que los aspirantes a biólogos moleculares debían aprender a ver la célula como una fábrica que contiene muchas cadenas de montaje entrelazadas de máquinas proteicas. En relación con el uso del término «máquina» en este contexto, Alberts ofreció la siguiente explicación:
«¿Por qué llamamos máquinas proteicas a los grandes ensamblajes proteicos que subyacen a la función celular? Precisamente porque, al igual que las máquinas inventadas por los seres humanos para lidiar de manera eficiente con el mundo macroscópico, estos ensamblajes proteicos contienen partes móviles altamente coordinadas. Dentro de cada ensamblaje proteico, las colisiones intermoleculares no solo se limitan a un pequeño conjunto de posibilidades, sino que la reacción C depende de la reacción B, que a su vez depende de la reacción A —tal y como ocurriría en una máquina de nuestra experiencia cotidiana.» (Alberts, 1998, p. 291)
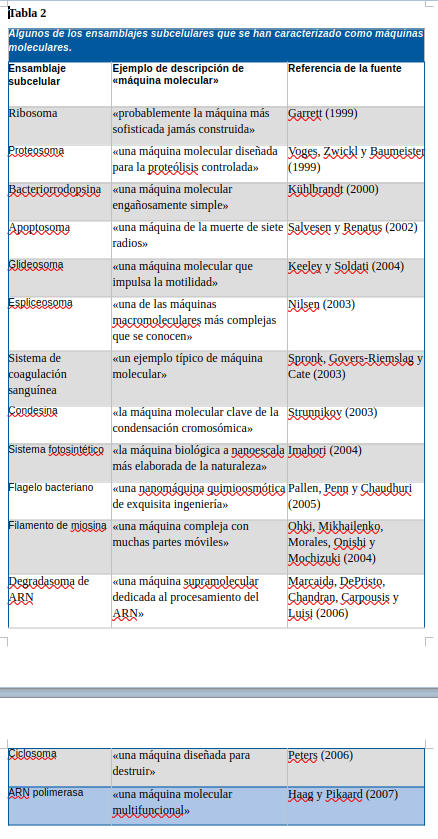
Esta terminología de las máquinas se popularizó rápidamente en la comunidad de la biología molecular y, año tras año, la frecuencia del término «máquina molecular» ha aumentado de forma constante en las revistas y publicaciones científicas, donde se ha convertido en una expresión de moda para describir prácticamente cualquier ensamblaje molecular altamente organizado y funcionalmente especializado de la célula. La Tabla 2 enumera una selección de las estructuras subcelulares que los biólogos han descrito recientemente como «máquinas moleculares».
A pesar de la popularidad del término «máquina molecular», un análisis bibliográfico revela que esta expresión aparece principalmente en artículos de revisión y solo en contadas ocasiones en trabajos de investigación. Cuando sí aparece en un trabajo de investigación, casi siempre lo hace en el título, el resumen o la introducción, y no en las partes del trabajo que describen realmente la investigación llevada a cabo y la interpretación de los resultados. Lo que esto sugiere es que, en lugar de desempeñar un papel teórico en la explicación de los ensamblajes subcelulares, el término «máquina molecular» se utiliza principalmente con fines retóricos, como una forma de introducir a los recién llegados al subcampo de la biología molecular dedicado a la caracterización estructural de grandes ensamblajes subcelulares. De hecho, la expresión «máquinas moleculares» se ha convertido en un título habitual para conferencias y sesiones académicas, por lo que parece funcionar principalmente a nivel social, al ayudar a individualizar un subconjunto concreto dentro de la comunidad de la biología molecular.
Sin embargo, las consecuencias negativas derivadas de la adopción del término «máquina molecular» superan con creces sus posibles beneficios pedagógicos y sociológicos. Esto se debe a que, gracias al uso persistente de este término por parte de los biólogos, los creacionistas actuales han encontrado precisamente el tipo de munición retórica que necesitan para revestir su creencia en un ser sobrenatural de una apariencia de respetabilidad científica. De hecho, el movimiento creacionista conocido como «diseño inteligente» (DI, en adelante) ha llegado a basarse en las llamadas «máquinas moleculares» de la biología molecular como principal fundamento empírico de sus afirmaciones sobre el diseño divino (véase Behe, 2001). Behe, uno de los principales defensores del DI, sostiene que muchos de los sistemas subcelulares altamente organizados que los biólogos moleculares describen como máquinas moleculares exhiben una «complejidad irreducible» que no podría haber evolucionado como resultado de causas naturales y que, por lo tanto, debe ser producto de una agencia inteligente. Behe (2006, p. 39) define un sistema irreduciblemente complejo como aquel compuesto por muchas partes que interactúan y contribuyen a la función del sistema, y en el que la eliminación de cualquiera de las partes provoca necesariamente que el sistema deje de funcionar. Ilustra este concepto utilizando una máquina, la trampa para ratones, que requiere la presencia simultánea de un resorte, una barra, una plataforma y otros componentes para atrapar ratones. Behe señala que la eficiencia de la trampa para ratones no aumenta gradualmente con la sucesiva adición de componentes. Más bien, todos los componentes deben estar en su lugar para que la máquina funcione en absoluto. Del mismo modo, Behe sostiene que las máquinas moleculares, como el flagelo bacteriano y el sistema de coagulación de la sangre, también constituyen sistemas irreduciblemente complejos.
El papel que desempeña la noción de máquina molecular en la defensa del DI de Behe es crucial, ya que proporciona una forma tácita de eludir la parte inductiva del Argumento del Diseño con el fin de establecer, sobre bases puramente analíticas, que, dado que las máquinas tienen diseñadores y que los sistemas vivos son conjuntos de máquinas moleculares, se deduce lógicamente que los sistemas vivos deben tener un diseñador. Como demostró Hume en sus Diálogos sobre la religión natural, el problema de este argumento no radica en inferir el diseño (y, por tanto, un diseñador) a partir de una máquina, sino en concebir ontológicamente los sistemas vivos como máquinas en primer lugar. Behe es muy consciente de esto, por lo que dedica tanta atención en sus escritos a enfatizar la naturaleza similar a una máquina de las células. Afortunadamente para Behe, esta tarea se ve enormemente facilitada por el hecho de que los biólogos moleculares son los primeros en utilizar lenguaje de máquinas en sus propias descripciones de los complejos subcelulares, como ilustra la Tabla 2. Por ello, Behe (2006, p. 218) puede afirmar que «la crítica de Hume al argumento del diseño, que sostiene una diferencia fundamental entre los sistemas mecánicos y los sistemas vivos, ha quedado obsoleta, refutada por el avance de la ciencia que ha descubierto la maquinaria de la vida». Behe se convierte en un mecanicista de facto cuando se trata de la célula, y es aquí donde la terminología de las máquinas moleculares, adoptada por los propios biólogos moleculares, resulta tan conveniente. De hecho, Behe aprovecha cualquier oportunidad para emplearla en sus escritos, como ilustra el siguiente pasaje:
«Los resultados acumulados [de la biología molecular] muestran con penetrante claridad que la vida se basa en máquinas —¡máquinas hechas de moléculas! Las máquinas moleculares transportan carga de un lugar a otro de la célula a lo largo de “autopistas” formadas por otras moléculas, mientras que otras actúan como cables, cuerdas y poleas para mantener la forma de la célula. Las máquinas activan y desactivan interruptores celulares, a veces matando a la célula o provocando su crecimiento. Las máquinas alimentadas por energía solar capturan la energía de los fotones y la almacenan en sustancias químicas. Las máquinas eléctricas permiten que la corriente fluya a través de los nervios. Las máquinas de fabricación construyen otras máquinas moleculares, así como a sí mismas. Las células nadan utilizando máquinas, se replican con maquinaria e ingieren alimento con maquinaria. En resumen, máquinas moleculares altamente sofisticadas controlan todos los procesos celulares.» (Behe, 2006, pp. 4–5)
En manos de Behe, el término «máquina molecular» ya no es una inofensiva figura retórica, sino una caracterización estrictamente literal de la naturaleza de los componentes celulares: «literalmente, hay máquinas reales dentro de las células de todos y así es como las denominan todos los biólogos que trabajan en este campo: máquinas moleculares» (Behe, 2005). La ingeniosidad del argumento de Behe a favor del DI reside, por tanto, en su hábil explotación de un término que los biólogos moleculares utilizan de manera imprecisa con fines retóricos, de tal forma que se aprovecha su autoridad como científicos para socavar sus propias afirmaciones. La consecuencia interesante de esto es que las refutaciones del DI formuladas por biólogos y filósofos se han reducido en gran medida a argumentos contra el OCM. Por ejemplo, en su crítica a Behe, Shanks y Joplin (1999, p. 281) señalan que «los sistemas biológicos reales son muy diferentes de los artefactos de ingeniería diseñados de forma económica, como las trampas para ratones. El argumento de [Behe] contra la evolución es, de hecho, un buen ejemplo de los peligros de quedar “atrapado” por una metáfora», con lo que, por supuesto, se refieren a la metáfora de la máquina. De manera similar, en su crítica al DI, Scott y Matzke (2007, p. 292) sostienen que «las diferencias entre los fenómenos biológicos y las máquinas construidas por el ser humano superan con creces las similitudes superficiales». En consecuencia, en un editorial titulado «Defendamos la evolución», Raff (2005, p. 274) hace hincapié en la necesidad de que los biólogos eviten el OCM en su enseñanza y sus escritos: « no caigamos en la trampa de los propagandistas del DI. […] Llamar a las células “máquinas que hacen X” o describir las estructuras biológicas como “bien diseñadas para hacer Y” será debidamente citado en la propaganda del DI como un biólogo más que apoya el diseño». Por lo tanto, queda claro que las consecuencias perniciosas que se derivan del uso retórico del OCM por parte de los biólogos eclipsan totalmente su potencial valor didáctico y sociológico en campos concretos como la biología molecular.
*
Ahora estamos en condiciones de responder a la pregunta con la que comenzamos esta sección. Los numerosos problemas del OCM destacados en la sección 3 no nos obligan a prescindir por completo del OCM. Sin embargo, sí exigen que se delimite adecuadamente el papel del OCM en la biología. Esto se ha logrado considerando de forma independiente las funciones teóricas, heurísticas y retóricas del OCM. He argumentado que, aunque el OCM es inadecuado como teoría del organismo, sigue siendo una valiosa herramienta heurística cuando se emplea en la investigación biológica. Abordar el estudio de los organismos como si fueran máquinas puede resultar muy provechoso y es, en cierta medida, necesario. No obstante, es crucial no permitir que la fecundidad empírica del OCM oscurezca el hecho innegable de que los organismos y las máquinas son fundamentalmente diferentes. El peligro de caer en una interpretación teórica del OCM está siempre presente, y por eso el uso retórico del OCM es tan peligroso. El precio que los biólogos deben pagar por su provechoso uso heurístico del OCM es que deben mantener la sobriedad intelectual necesaria para resistir la tentación de sucumbir al atractivo teórico de esta noción intuitivamente convincente.
5. Conclusión
En este artículo he pretendido demostrar que el OCM es hoy en día uno de los obstáculos más graves que impiden un mayor avance en nuestra comprensión teórica de los sistemas vivos. En esencia, el OCM reduce sistemáticamente a los organismos a las características que es capaz de explicar y descarta todo lo demás. El éxito a la hora de explicar estas características seleccionadas contribuye, a su vez, a reforzar la conceptualización original del organismo como una máquina. Los aspectos más recalcitrantes del organismo que no encajan en el OCM se ignoran o se descartan por considerarlos sin importancia. Lamentablemente, lo que se descuida en el OCM es precisamente aquello que hace que los organismos sean más distintivos, a saber, su finalidad intrínseca y su organización autónoma. No debería sorprender que los conceptos típicamente asociados a estas dos características no mecánicas, a saber, la teleología y el holismo, sean generalmente tratados con escepticismo y recelo por la comunidad científica. Dado que el edificio de la ciencia moderna se construyó históricamente sobre cimientos mecanicistas, es de esperar que lo que queda fuera del alcance del mecanicismo haya tendido a descartarse fácilmente como místico o anticientífico. Como afirma Rosen (Rosen, 1991, pp. xv–xvi), «durante los últimos tres siglos, las ideas de mecanismo y máquina han constituido la esencia misma del adjetivo “científico”; por lo tanto, rechazarlas parece un rechazo de la ciencia misma». Pero esto no es más que un prejuicio. Una vez que se deja atrás y se comprende que el mecanicismo, a pesar de su importancia histórica, no predetermina ni agota el significado de la ciencia, la teoría biológica se libera de la necesidad de ajustarse al OCM y es capaz de explorar modelos conceptuales alternativos que intentan afrontar la complejidad del organismo en sus propios términos.
Agradecimientos
Doy las gracias a Peter McLaughlin, Paul Griffiths, Lenny Moss, John Dupré, Philippe Huneman, Arnon Levy y Dan McShea por sus útiles debates y comentarios constructivos sobre las primeras versiones y/o borradores recientes de este artículo. También agradezco al público asistente a la Conferencia Internacional sobre Vida Artificial en Winchester, al Encuentro Europeo de Doctorandos en Filosofía de las Ciencias de la Vida en Rovigo y al Taller sobre Perspectivas Históricas y Filosóficas del Concepto de Organismo en París, por sus comentarios sobre las presentaciones relacionadas con este tema. Deseo agradecer especialmente a Russell Powell por su invitación a colaborar en este número especial. Por último, me complace agradecer el apoyo financiero del Instituto Konrad Lorenz para la Investigación en Evolución y Cognición, así como del Instituto Cohn para la Historia y la Filosofía de la Ciencia y las Ideas.
Estudios de Historia y Filosofía de las Ciencias Biológicas y Biomédicas n.º 44, diciembre de 2013.
Notas:
[8] La historia de la biología ofrece un ejemplo elocuente de este punto. En el siglo XVIII, los fisiólogos de la Académie des Sciences permitieron que el ingeniero Vaucanson influyera en los debates sobre la naturaleza de la digestión gracias a su creación de un pato mecánico que parecía capaz de digerir los alimentos, ya que comía granos y luego los excretaba, imitando así las entradas y salidas conductuales de un pato vivo. Tras la muerte de Vaucanson, un examen minucioso del mecanismo de deglución del pato reveló que la ingesta de grano y la excreción no guardaban relación, y que la parte trasera del pato había sido cargada con excrementos falsos antes de cada actuación (véase Riskin, 2003).
[9] Como señaló Cossa en La cybernétique (1957), «lo que es inherente al ser vivo no es meramente el medio, sino el fin mismo: la preservación de la vida, la preservación de la continuidad de la existencia mediante la adaptación al entorno. No hay nada parecido en el homeostato, este no tiene un objetivo último inherente. Si un ser vivo, cuyo equilibrio se ha visto alterado, prueba perseverantemente, una tras otra, todas las formas posibles de adaptarse a su nuevo entorno, esto se explica como un esfuerzo por sobrevivir. Si el homeostato prueba sus 390 625 combinaciones una tras otra, solo lo hace porque eso es lo que Ashby quería de él» (citado en Oparin, 1961, p. 26)
[10] El ampliamente difundido informe de los investigadores del Instituto J. Craig Venter (Gibson et al., 2010), que algunos aclamaron como la creación de vida artificial, consistió en realidad en el trasplante de una copia sintética de un genoma natural a una célula preexistente a la que se le había eliminado su propio genoma. Aunque sin duda se trata de una hazaña tecnológica notable, tal logro aún está lejos de la síntesis artificial de una célula viva.
[11] En palabras de Oparin (1961, p. 19), la «identificación de los seres vivos con máquinas se consideraba [históricamente] como la única forma de salvar a la ciencia de la entelequia mística de los vitalistas, el puente que nos llevaría de la física y la química a la biología».
[12] Es por esta razón que Driesch recurrió a las propiedades holísticas de los organismos en una fase temprana de su desarrollo para demostrar la insuficiencia del OCM y formular su «primera prueba del vitalismo» (véase Driesch, 1908, pp. 118-149).
Referencias:
Véase: https://sniadecky.wordpress.com/2026/05/09/nicholson-organisms-machines-en/
———————