Por Ekaterina Dadachova, Arturo Casadevall
RESUMEN DE AVANCES RECIENTES
La vida en la Tierra siempre ha existido bajo el influjo de la radiación ionizante. Sin embargo, los hongos parecen interactuar con la radiación ionizante de manera diferente a otros habitantes de la Tierra. Datos recientes muestran que las especies de hongos melanizados, como los del reactor de Chernóbil, responden a la radiación ionizante con un crecimiento acentuado. Los hongos colonizan las estaciones espaciales y se adaptan morfológicamente a condiciones extremas. La exposición a la radiación provoca la regulación al alza de muchos genes clave, y una vía de recombinación inducible mediada por microhomología podría ser un mecanismo potencial de evolución adaptativa en eucariotas. El descubrimiento de organismos melanizados en entornos de alta radiación, las estaciones espaciales, las montañas antárticas y en el agua de refrigeración de los reactores, combinado con el fenómeno del «radiotropismo», plantea la tentadora posibilidad de que las melaninas tengan funciones análogas a otros pigmentos captadores de energía, como las clorofilas.
INTRODUCCIÓN
La vida surgió en la Tierra en una época en la que la radiación de fondo era mucho mayor y las primeras formas de vida debían tener una considerable resistencia a la radiación. Aunque los niveles actuales de radiación de fondo son mucho más bajos que en los inicios de la Tierra, la vida terrestre sigue existiendo en un campo de radiación. Por ejemplo, el 90 % de la dosis de radiación anual de una persona que vive en EE. UU. procede de fuentes naturales como la radiación cósmica y las rocas radiactivas (1.). Sin embargo, existen pruebas considerables de que los hongos responden a la radiación de una manera que puede diferir de otras formas de vida. Se han encontrado grandes cantidades de esporas fúngicas altamente melanizadas en depósitos del período Cretácico Inferior, cuando se extinguieron muchas especies de animales y plantas. Este período coincide con el paso de la Tierra por el «cero magnético», lo que provocó la pérdida de su «escudo» contra la radiación cósmica (2). Además, se ha sugerido que la radiación de una supuesta estrella fugaz llamada Némesis podría haber contribuido a los eventos de extinción (3). Los hongos en general, y especialmente los melanizados, son altamente radioresistentes cuando se someten a altas dosis de radiación ionizante en condiciones experimentales (4) – (7). Es comprensible que estas capacidades inusuales de los eucariotas para sobrevivir o incluso beneficiarse de la exposición a la radiación ionizante contrasten con la opinión general de que la radiación es uniformemente perjudicial para la vida. El tema de las interacciones de las células fúngicas con los radionucleidos es de considerable interés para la reparación ambiental (revisado en [8]), pero ese fenómeno es de naturaleza química y es diferente de las interacciones con la radiación ionizante. En consecuencia, en esta revisión nos centraremos en los hallazgos recientes sobre la interacción de los hongos con la radiación externa, como los hongos que residen en entornos altamente radiactivos, el radiotropismo de los hongos asociados a Chernóbil, los hongos en el espacio, los primeros intentos de descifrar el mecanismo de utilización de la energía de la radiación por parte de los hongos, así como algunas ideas sobre los efectos genéticos de la radiación en los hongos.
HONGOS QUE HABITAN EN ENTORNOS CON ALTOS NIVELES DE RADIACIÓN
Los hongos melanizados habitan en algunos entornos extraordinariamente extremos del planeta, como las regiones del Ártico y la Antártida y los terrenos de gran altitud, caracterizándose estos últimos hábitats por los niveles de radiación naturalmente más altos que en las altitudes más bajas (9). El «Cañón de la Evolución» en Israel es un sitio popular para el estudio de la adaptación de los organismos a su entorno. Tiene dos vertientes: la vertiente «europea», orientada al norte, y la vertiente «africana», orientada al sur, esta última recibe entre un 200 y un 800 % más de radiación solar que la vertiente norte y está poblada por muchas especies de hongos melanizados como el Aspergillus niger, que contiene tres veces más melanina que la misma especie de la vertiente orientada al norte (10). Curiosamente, cuando especies de Alternaria, Aspergillus, Humicola, Oidiodendron y Staphylotrichum de ambas laderas se sometieron a altas dosis (hasta 4000 Gy) de radiación de Co-60, los aislados de la ladera sur crecieron a mayor velocidad que los aislados de la ladera norte (11).
Entre los entornos con alta radiación resultante de actividades humanas, destacan dos ejemplos. En primer lugar, las especies de hongos melanizados colonizan las paredes del reactor dañado de Chernóbil, donde están expuestos a un campo de radiación constante y alta (12). En segundo lugar, se encuentran especies de hongos melanizados en el agua de la llamada piscina de refrigeración del reactor. Esta agua circula por el núcleo del reactor nuclear con fines de refrigeración y es extremadamente radiactiva. Estas piscinas contienen grandes cantidades de hongos, cocos, bacilos grampositivos y algunos bacilos gramnegativos. El análisis de esta microflora del agua del reactor ha llevado a sugerir que los altos flujos de radiación seleccionan tipos de microorganismos altamente radioresistentes, que manifiestan aumentos en las actividades de catalasa y nucleasa (13).
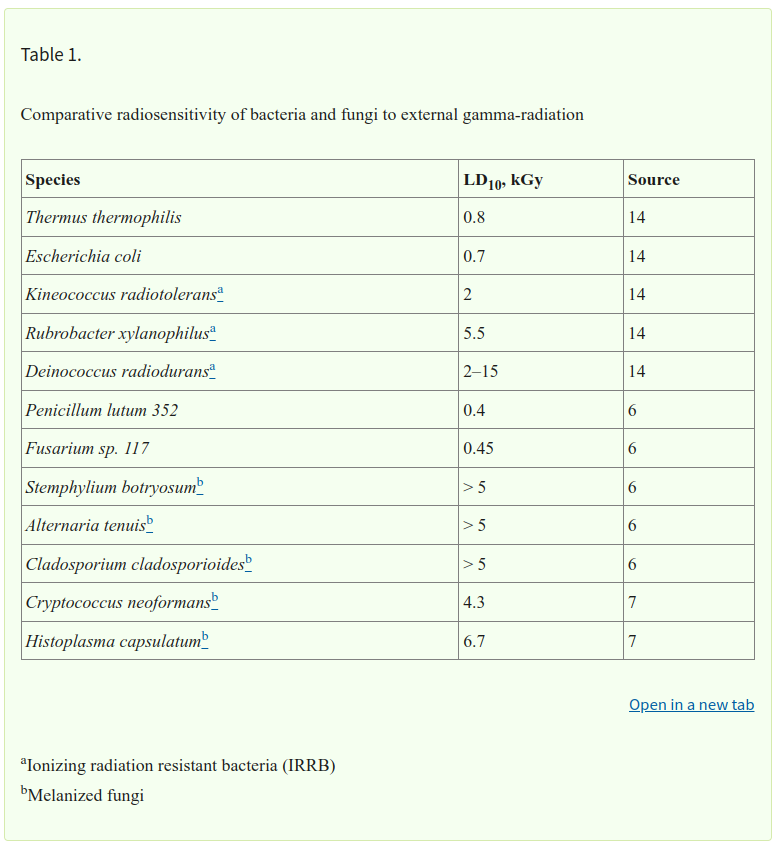
RADIOSENSIBILIDAD COMPARATIVA DE BACTERIAS Y HONGOS
La bacteria Deinococcus radiodurans se considera el microorganismo más resistente a la radiación conocido, con una DL10 para algunas cepas que se acerca a los 15 kGy (14). La dosis estándar para la irradiación de alimentos en EE. UU. es de 1 kGy, que se considera suficiente para matar la mayor parte de los microorganismos contaminantes de los alimentos, ya que solo unas pocas cepas de bacterias tienen valores de DL10 superiores a 1 kGy (Tabla 1). Estas bacterias se denominan bacterias resistentes a la radiación ionizante (IRRB) (14). Sin embargo, muchos hongos, especialmente los melanizados, son muy radiorresistentes, con valores de DL10 que se acercan o superan 1 kGy (Tabla 1). Esta radiorresistencia de los hongos no es muy conocida y debe tenerse en cuenta cuando se utiliza radiación gamma para la esterilización de alimentos o suministros médicos.
RADIOTROPISMO DE LOS HONGOS ASOCIADOS A CHERNÓBIL
Zhdanova et al. informaron de que algunos de los hongos que crecían en la zona alrededor del lugar del accidente nuclear de Chernóbil de 1986 tenían la capacidad de crecer y descomponerse en las llamadas «partículas calientes», trozos de grafito del reactor n.º 4 destruido que están contaminados con varios radionucleidos de larga vida (15, 16). Denominaron «radiotropismo» a esta atracción de los hongos por la radiación. En su trabajo más reciente, excluyeron los posibles efectos de interferencia del carbono en el crecimiento direccional de los hongos exponiéndolos a los haces de radiación colimados externos de los radionucleidos P-32 y Cd-109, que son emisores beta y gamma, respectivamente (17). Los autores midieron el «ángulo de retorno», que definieron como el ángulo entre el punto de impacto de la radiactividad en el recipiente de cultivo y la dirección de crecimiento de la porción distal de la hifa emergente de cada espora. Un ángulo de retorno bajo (<90°) indica un crecimiento medio de la hifa hacia la fuente de radiactividad y un ángulo alto (90-180°) indica un crecimiento en dirección opuesta a la fuente. Los hongos utilizados en el experimento se aislaron de las zonas contaminadas de Chernóbil, o se aislaron antes de la explosión o de los sitios remotos. En total, se investigaron 27 respuestas de interacciones entre aislados de hongos y la fuente de radiación. De estos, 18 (66,7 %) mostraron estimulación positiva del crecimiento hacia la fuente de radiación (ángulo de retorno medio bajo), y ocho no mostraron respuesta. En la figura 1 se muestran ejemplos de resultados que muestran el «ángulo de retorno» medio. Se observó un crecimiento dirigido estadísticamente significativo hacia la fuente de radiación de Cd-109 para Penicillium roseopurpureum 147 (de suelo contaminado de Red Forest), P. hirsutum 3 (partículas calientes), aislados de Cladosporium cladosporioides 60 y 10 (de la sala del reactor del cuarto bloque). C. sphaerospermum 3176, aunque aislado de suelo de control no contaminado, también mostró una respuesta positiva. Se observó una tendencia hacia el crecimiento direccional, aunque no estadísticamente significativa, para C. cladosporioides 396 y Paecilomyces lilacinus 101 (ambos aislados de suelos no contaminados) y para Penicillium lanosum (del cuarto bloque) y Paecilomyces lilacinus 1941 (suelo de Red Forest), ambos aislados originalmente de zonas con altos niveles de radiación. Los autores concluyeron que tanto la radiación beta como la gamma promovían el crecimiento direccional de hongos desde áreas contaminadas y limpias hacia las fuentes de radiación ionizante.
En su trabajo posterior, publicado en 2006-2007, el mismo grupo investigó la influencia de la radiación externa del Sn-121 (emisor gamma de baja energía) y el Cs-137 (emisor mixto beta y gamma de alta energía) no solo en el crecimiento hifal de los hongos de las regiones de Chernóbil contaminadas radiactivamente frente a los controles, sino también en la germinación de sus esporas (18, 19). Observaron que la radiación promovía la germinación de las esporas en especies de regiones contaminadas, lo que denominaron «radiestimulación». En contra de sus resultados anteriores (17), observaron la «radiestimulación» solo en las especies de regiones contaminadas, pero no en los aislados de las zonas limpias. Llamaron a este fenómeno «respuesta radioadaptativa». También observaron los mismos resultados en las respuestas de los hongos de las zonas contaminadas a la luz ([20]). Sin embargo, aunque es muy probable que los hongos expuestos a largo plazo a niveles elevados de radiación tengan propiedades adaptativas, las limitaciones del trabajo experimental descrito en ([18]–[20]) podrían interferir en la capacidad de los autores para observar también la radiostimulación en los hongos de las zonas limpias. Por ejemplo, la actividad de las fuentes radiactivas utilizadas en los últimos estudios (del 18 al 20) fue aproximadamente 1000 veces menor que la utilizada en trabajos anteriores (del 17), lo que podría haber sido insuficiente para promover el crecimiento de las hifas. Este podría haber sido el caso especialmente para el Sn-121 de baja energía; además, las partículas beta del Cs-137 podrían haber sido absorbidas por el material de la placa de Petri con los hongos, ya que el haz colimado provenía de abajo y, por lo tanto, no han contribuido a las dosis de radiación reales que podrían haber sido sobrestimadas.
HONGOS QUE HABITAN EN LA NAVE ESPACIAL
Otro entorno de alta radiación al que se han adaptado los hongos es la órbita de las naves espaciales. El análisis de la atmósfera de la estación orbital rusa Mir reveló la presencia ubicua de muchos microorganismos (21). Las posibles fuentes de contaminación de la estación espacial son los materiales de vuelo durante la fabricación y el montaje, la entrega de suministros a la estación espacial, los propios suministros y la contaminación secundaria de la tripulación y cualquier otro material biológico a bordo, por ejemplo, animales, plantas y microorganismos utilizados en experimentos científicos (21). La contaminación fúngica supone ciertas amenazas para el bienestar de la tripulación, no solo porque algunos de esos hongos son patógenos humanos potenciales, sino también porque los hongos poseen potentes sistemas enzimáticos y secretan diversos metabolitos capaces de degradar materiales estructurales dentro de la nave espacial, desde polímeros hasta diversas aleaciones.
El estudio de la contaminación ambiental a bordo de la Estación Espacial Internacional (ISS) reveló la presencia de muchas especies de hongos en las superficies y en el aire (Tabla 2), siendo Aspergillus sp., Penicillium sp. y Saccharomyces sp. los géneros más dominantes entre los hongos. Se recuperó una población diversa de Aspergillus (13 especies), mientras que la diversidad fue menos pronunciada en el caso de Penicillium (5 especies) y Cladosporium (4 especies) (p. 22). Los niveles de radiación ionizante que estos hongos encuentran en las estaciones espaciales, aproximadamente 4 cGy por año (p. 23), no son fungicidas (p. 4–7, p. 13) y permiten que los hongos prosperen siempre que los niveles de humedad sean suficientes. Curiosamente, se descubrió que muchos de los microorganismos que habitan en la estación espacial, tanto bacterias como hongos, estaban pigmentados o melanizados, lo que sugiere la utilidad de la presencia de pigmentos en esas células en condiciones extremas.

Otro aspecto importante de la microbiología relacionada con los vuelos espaciales es la posibilidad de que los microorganismos que habitan en las naves espaciales cambien sus propiedades hasta tal punto que se vuelvan peligrosos para los habitantes de la Tierra cuando la nave regrese a la Tierra. Lo más probable es que esos microorganismos se encuentren en las superficies exteriores de la nave, donde estarían expuestos a las condiciones extremas del espacio abierto. Para investigar esa posibilidad, los investigadores llevaron a cabo el experimento «Biorisk» (n.º 24). La investigación por microscopía electrónica de Aspergillus versicolor y Penicillum expansum expuestos a condiciones de espacio abierto durante 7 meses reveló muchos cambios morfológicos, que aparentemente permitieron a esos hongos sobrevivir. Por ejemplo, la cápsula de polisacárido y la capa de melanina en P. expansum aumentaron significativamente en comparación con las muestras de control, así como el número de mitocondrias y vacuolas en los hongos expuestos al espacio fue mucho mayor que en los controles.

MECANISMO SUGERIDO DE UTILIZACIÓN DE LA ENERGÍA DE RADIACIÓN POR LOS HONGOS
Dada la resistencia y adaptabilidad de los hongos a los entornos de radiación ionizante y que muchos hongos producen melanina, planteamos la hipótesis de que la radiación podría cambiar las propiedades electrónicas de la melanina, de modo que el pigmento podría funcionar en la transducción de energía y que esto podría mejorar el crecimiento de los hongos melanizados. En apoyo de esta noción, la irradiación ionizante cambió la señal de resonancia de espín electrónico (ESR) de la melanina, en consonancia con los cambios en la estructura electrónica (25). La melanina irradiada manifestó un aumento de 4 veces en su capacidad para reducir el NADH en relación con la melanina no irradiada. El análisis por HPLC de la melanina de hongos cultivados en diferentes sustratos reveló complejidad química, dependencia de la composición de la melanina del sustrato de crecimiento y posible influencia de la composición de la melanina en su interacción con la radiación ionizante. La interacción con la radiación ionizante se estudió en tres especies de hongos: Cryptococcus neoformans, que puede crecer tanto en forma melanizada como no melanizada dependiendo de la presencia de sustrato exógeno, y dos especies intrínsecamente melanizadas, Wangiella dermatitidis y Cladosporium sphaerospermum, siendo esta última una de las especies predominantes que habitan en el reactor destruido de Chernóbil. Los ensayos XTT (hidróxido de 2,3-bis(2-metoxi-4-nitro-5-sulfofenil)-5-[(fenilamino)carbonil]-2H-tetrazolio) y MTT (bromuro de 2-(4,5-dimetil-2-tiazolil)-3,5-difenil-2H-tetrazolio) mostraron un aumento de la actividad metabólica de las células irradiadas y melanizadas de C. neoformans irradiadas en relación con las células no melanizadas irradiadas, lo que concuerda con la observación de que la exposición a la radiación ionizante mejora las propiedades de transferencia de electrones de la melanina. Las células melanizadas de W. dermatitidis y C. neoformans expuestas a una radiación ionizante aproximadamente 500 veces superior a la de fondo crecieron significativamente más rápido, como indican las UFC más altas, la biomasa de peso seco más elevada y la incorporación de acetato C-14 tres veces mayor que las células melanizadas no irradiadas o los mutantes albinos irradiados. Además, la radiación potenció el crecimiento de las células melanizadas de C. sphaerospermum en condiciones de nutrientes limitados. Las observaciones de que las células fúngicas melanizadas manifestaban un mayor crecimiento en relación con las células no melanizadas tras la exposición a radiación ionizante plantearon la intrigante posibilidad de que la melanina pueda funcionar en la captura y utilización de energía (25).
Con respecto a la posibilidad de que los hongos utilicen la radiación ionizante para obtener energía, es interesante observar la literatura más antigua que informa de la fijación de carbono por hongos en condiciones de nutrientes limitados (26)–(28). Se informó de que los hongos utilizan CO2 para la síntesis de los intermedios del ciclo del ácido tricarboxílico (TCA). La función biosintética del ciclo del TCA requiere un suministro constante de oxaloacetato, succinil-CoA y 2-oxoglutarato, y esas reacciones que reponen el suministro de los intermedios del ciclo del TCA se han denominado anapleróticas. Varias enzimas han sido relacionadas con la fijación anaplerótica de CO2 por microorganismos y la mayoría de los estudios especifican piruvato y fosfoenolpiruvato carboxilasas y fosfoenolpiruvato carboxiquinasa como las principales actividades. Esta fijación de CO2 se produce en luz blanca y conduce a un aumento de la biomasa, a diferencia de la fijación en la oscuridad como parte de la gluconeogénesis, que no conduce a una ganancia neta de carbono. Es tentador sugerir que, en condiciones de nutrientes limitados, los hongos melanizados podrían utilizar este mecanismo de fijación de CO 2 mediante la transducción por la energía de la melanina de la radiación ionizante en lugar de la luz blanca, y quizás esto debería probarse experimentalmente en futuros trabajos.
Aparte de su papel en la transducción de energía, la melanina parece tener importantes propiedades radioprotectoras. C. neoformans y Histoplasma capsulatum no melanizados son muy resistentes a la radiación, pero la presencia de melanina aumenta aún más la supervivencia a dosis más altas. La percepción actual de las propiedades radioprotectoras de la melanina es que apaga los radicales libres citotóxicos de corta duración y, por tanto, previene el daño en el ADN. Sin embargo, también planteamos la hipótesis de que las propiedades radioprotectoras de la melanina en los microorganismos son el resultado de una combinación de protección física y extinción de los radicales libres citotóxicos. Cuando se fragmentaron los restos de melanina aislados de las células melanizadas, estos perdieron gran parte de sus propiedades de protección radiológica, lo que indica que la disposición esférica de las partículas de melanina en la cáscara hueca contribuye a la protección radiológica. Llegamos a la conclusión de que la melanina protege a los hongos contra la radiación ionizante y que sus propiedades radioprotectoras dependen de su composición química, de la extinción de los radicales libres y de su disposición espacial esférica (29).
EFECTOS GENÉTICOS DE LA RADIACIÓN EN LOS HONGOS
Se pueden obtener algunas ideas sobre los efectos genéticos de la radiación ionizante en los hongos a partir de los estudios realizados con S. cerevisiae. Kimura et al. utilizaron microarrays de ADN para investigar un perfil de expresión génica posterior a la irradiación en células de levadura expuestas a rayos X y rayos gamma (30). El análisis de microarrays reveló que tanto los rayos X como los rayos gamma regulaban al alza los genes relacionados con el ciclo celular y el procesamiento del ADN, la defensa y la virulencia de reparación celular, el destino de las proteínas y las células, y el metabolismo (Fig. 2). Del mismo modo, para ambos tipos de rayos, los genes regulados negativamente pertenecían en su mayoría a las categorías de transcripción y síntesis de proteínas, ciclo celular y procesamiento de ADN, control de la organización celular, destino celular y metabolismo de compuestos C y carbohidratos. Los cambios en los cultivos irradiados con rayos gamma se observaron más tarde que en los irradiados con rayos X. Los autores atribuyeron las diferencias en el curso temporal a las diferencias en la transferencia de energía lineal entre los rayos X de baja energía y los rayos gamma de alta energía. Bennett et al. investigaron qué genes de S. cerevisiae son realmente responsables de la resistencia a la radiación ionizante y descubrieron que muchos de estos genes eran responsables de funciones tan importantes como la reparación (RAD50, RAD51), la recombinación (HRP1), la estabilidad cromosómica (CHL1, CTF4), la endocitosis (VID21), la degradación de la ubiquitina (GRR1), la transcripción (BUR2) y algunas otras (31). Un estudio de Ustilago maydis, también conocido por su extrema resistencia a la radiación, reveló un conjunto similar de genes (32). Los autores concluyeron que la supervivencia de U. maydis tras la exposición a altas dosis de radiación es el resultado de los niveles/acciones de las proteínas implicadas en la reparación del ADN, más que de la presencia de un sistema de recombinación especializado como el de D. radiodurans, y que la naturaleza biotrófica de U. maydis condujo a la aparición de un sistema eficiente de reparación del ADN (32). Curiosamente, muchos de los genes de resistencia a la radiación comparten una homología significativa con los genes humanos que podría aprovecharse en el desarrollo de nuevas terapias contra el cáncer basadas en la radiación.
La radiación ionizante genera roturas de una sola hebra (SSB), roturas de doble hebra (DSB), daños en las bases y entrecruzamientos de ADN. Las células eucariotas reparan las DSB mediante dos mecanismos, siendo el primero la recombinación homóloga. El segundo mecanismo se denomina recombinación ilegítima, o unión de extremos no homólogos (NHEJ), e implica la unión de extremos en ausencia de homología de secuencia de ADN (33). Algunos eventos de recombinación ilegítima se caracterizan por unos pocos pares de bases (pb) de homología compartidos en los extremos de las dos uniones de recombinación, lo que se denomina recombinación mediada por microhomología (MHMR) (34). Chan et al. estudiaron la MHMR en S. cerevisiae irradiada con 50 Gy de rayos gamma (35) y demostraron que una vía MHMR en todo el genoma inducida por DSB podría conducir a reordenamientos genómicos a gran escala después de que un único extremo de DSB invada otra ubicación genómica. Tal fenómeno puede proporcionar beneficios para desarrollar variantes genéticas que tengan ventajas de crecimiento bajo estrés genotóxico. Concluyeron que la vía MHMR inducible podría ser un mecanismo potencial de evolución adaptativa en eucariotas. Estas observaciones podrían explicar la respuesta radioadaptativa en hongos descrita por el grupo de Zhdanova (18.20), pero son una explicación poco probable para los efectos de crecimiento incrementado de los organismos melanizados irradiados, que respondieron en cuestión de horas.
MELANINAS Y RADIACIÓN EN PERSPECTIVA
Los pigmentos de melanina se encuentran en todos los reinos biológicos, lo que sugiere que estos compuestos son moléculas antiguas que surgieron al principio de la evolución. Las melaninas son polímeros complejos con una variedad de propiedades que pueden producirse enzimáticamente a partir de precursores relativamente simples. Un aspecto notable de las melaninas es su capacidad para absorber todo tipo de radiación electromagnética (36), lo que les confiere la capacidad tanto de transducción como de protección energética. Los hallazgos de organismos melanizados en entornos de alta radiación, como el reactor dañado de Chernóbil, la estación espacial, las montañas antárticas y el agua de refrigeración de reactores, combinados con el fenómeno del «radiotropismo», plantean la tentadora posibilidad de que las melaninas tengan funciones análogas a otros pigmentos de captación de energía, como las clorofilas.
Agradecimientos
E. Dadachova cuenta con el apoyo de la subvención AI60507 del Instituto Nacional de Alergias y Enfermedades Infecciosas (NIAID); A. Casadevall, con las subvenciones AI033142 y AI033774 del NIAID.
Referencias:
En el artículo original: https://pmc.ncbi.nlm.nih.gov/articles/PMC2677413/
———————