Charlotte Kuperwasser y Wafik S. El-Deiry, enero de 2026

RESUMEN
Un número creciente de publicaciones revisadas por pares han informado de diversos tipos de cáncer que aparecen en asociación temporal con la vacunación o la infección por COVID-19. Para caracterizar la naturaleza y el alcance de estos informes, se llevó a cabo una búsqueda bibliográfica sistemática desde enero de 2020 hasta octubre de 2025, basándose en criterios de elegibilidad específicos. Un total de 69 publicaciones cumplieron los criterios de inclusión: 66 informes a nivel de artículo que describían 333 pacientes en 27 países, 2 investigaciones retrospectivas a nivel de población (Italia: cohorte de ~300 000 personas, y Corea: cohorte de ~8,4 millones de personas) que cuantificaban las tendencias de incidencia y mortalidad por cáncer entre las poblaciones vacunadas, y un análisis longitudinal de ~1,3 millones de miembros del ejército estadounidense que abarcaba los periodos previos y posteriores a la pandemia. La mayoría de los estudios documentaron neoplasias hematológicas (linfomas no Hodgkin, linfomas cutáneos, leucemias), tumores sólidos (mama, pulmón, melanoma, sarcoma, cáncer de páncreas y glioblastoma) y cánceres asociados a virus (carcinoma de Kaposi y de células de Merkel). En todos los informes surgieron varios temas recurrentes: (1) progresión inusualmente rápida, recurrencia o reactivación de una enfermedad indolente o controlada preexistente, (2) hallazgos histopatológicos atípicos o localizados, incluida la afectación de los lugares de inyección de la vacuna o los ganglios linfáticos regionales, y (3) posibles vínculos inmunológicos entre la infección aguda o la vacunación y la latencia tumoral, el escape inmunológico o los cambios en el microambiente. El predominio de las observaciones a nivel de casos y los datos preliminares a nivel de población demuestran que se encuentra en una fase temprana de detección de posibles señales de seguridad. Estos hallazgos subrayan la necesidad de realizar estudios epidemiológicos, longitudinales, clínicos, histopatológicos, forenses y mecanicistas rigurosos para evaluar si la vacunación o la infección por COVID-19 pueden estar relacionadas con el cáncer y en qué condiciones.
Introducción
La pandemia de COVID-19 y el despliegue generalizado de nuevas vacunas basadas en ARNm y vectores virales han transformado el panorama de la inmunología humana [1–4]. Nunca antes una proporción tan grande de la población mundial había estado expuesta simultáneamente a inmunógenos basados en ácidos nucleicos, sistemas de administración de nanopartículas lipídicas (LNP) y regímenes de refuerzo repetidos durante un período relativamente corto. La escala sin precedentes que se movilizó en respuesta a la pandemia de COVID-19 ha generado y sigue generando una gran cantidad de datos clínicos, moleculares y epidemiológicos, que revelan respuestas biológicas que van más allá de la activación y las respuestas inmunitarias tradicionales inducidas por las vacunas. Entre ellas se incluyen una serie de síndromes neurológicos, autoinmunitarios e inflamatorios posteriores a la infección y a la vacunación, como miocarditis, neuropatías mediadas por el sistema inmunitario, citopenias autoinmunitarias, respuestas inflamatorias sistémicas [5–7], así como la coexistencia temporal con diagnósticos de cáncer, recidivas o trayectorias de la enfermedad inesperadamente rápidas [8–11]. Estos acontecimientos han dado lugar a una amplia investigación clínica y ponen de relieve la capacidad de la activación inmunitaria inducida por la vacuna para perturbar la homeostasis inmunitaria en personas susceptibles. Es importante destacar que muchas de estas afecciones se caracterizan por una desregulación de las citoquinas, una alteración de la señalización inmunitaria innata y adaptativa y respuestas inflamatorias específicas de los tejidos, vías que también están implicadas en la iniciación y progresión de los tumores y en la vigilancia inmunitaria. La presente revisión se centra específicamente en las observaciones relacionadas con el cáncer dentro de este contexto más amplio de perturbación inmunitaria tras la vacunación.
Casi seis años después de que se reconociera la pandemia a principios de 2020, la bibliografía mundial actual sobre la infección por COVID-19 o la vacunación y el cáncer sigue siendo escasa, heterogénea y limitada en gran medida a informes de casos y pequeñas series de casos, lo que resulta insuficiente para respaldar conclusiones definitivas sobre la causalidad o la cuantificación del riesgo. Los prospectos de las vacunas contra la COVID-19 publicados por la Administración de Alimentos y Medicamentos (FDA) [12–15] indican específicamente que no se ha evaluado su carcinogenicidad o genotoxicidad, ni se han estudiado tras múltiples dosis de vacunas y refuerzos o en combinación con una infección posterior por SARS-CoV-2.
Durante la pandemia de COVID, se predijo que las tasas de cáncer aumentarían durante y después de la COVID debido a la reducción de las pruebas de detección y del acceso al tratamiento durante la pandemia. Sin embargo, las tasas de cáncer entre las personas más jóvenes, por ejemplo, con cáncer de colon de aparición temprana, han ido aumentando durante dos décadas [16, 17]. Las tasas de colangiocarcinoma y cáncer de endometrio también han ido aumentando. Las muertes por cáncer superaron las 600 000 en Estados Unidos por primera vez en 2024 y se prevé que en 2025 también aumenten [18]. En el momento de redactar esta revisión, no hay estudios poblacionales publicados en Estados Unidos con un seguimiento de la mortalidad o la incidencia del cáncer más allá de los 42 días para comparar los resultados tras la infección por COVID con los de no haber tenido la infección o estar vacunado frente a no estarlo. Esto se debe en parte a la falta de bases de datos de buena calidad que contengan dicha información. Existe un Consorcio sobre la COVID y el Cáncer (CCC) financiado por el Instituto Nacional del Cáncer (NCI), pero no ha publicado nada específico sobre este tema.
En un contexto de evidencia clínica limitada y toxicología preclínica incompleta, un estudio reciente informó de que las vacunas de ARNm contra el SARS-CoV-2 podrían sensibilizar los tumores al bloqueo de los puntos de control inmunitarios [19], lo que ha dado lugar a una interpretación generalizada de que la vacunación con ARNm contra la COVID-19 podría potenciar las respuestas antitumorales en pacientes con melanoma o cáncer de pulmón no microcítico (CPNM) sometidos a inhibición de los puntos de control inmunitarios. Además, en el análisis, la vacunación con ARNm se asoció con un aumento de la señalización del interferón tipo I y una elevada expresión de PD-L1 en el tumor. Sin embargo, la regulación al alza de PD-L1 en ausencia de terapia con inhibidores de puntos de control se asocia generalmente con una mayor evasión inmunitaria del tumor y resistencia a la citotoxicidad mediada por células T, lo que plantea dudas sobre la interpretación biológica de estos hallazgos. Aunque las terapias basadas en interferón han demostrado su utilidad clínica en el melanoma, el estudio no proporcionó análisis comparativos entre el tratamiento con interferón y la combinación de la vacunación con ARNm con el bloqueo de puntos de control. Además, el estudio no abordó las limitaciones clave, las explicaciones mecánicas alternativas ni el contexto clínico más amplio necesario para interpretar plenamente los efectos descritos.
Esta ausencia de evaluación de las vacunas contra la COVID-19 en cuanto a carcinogenicidad o genotoxicidad motivó una revisión sistemática y una síntesis de las pruebas disponibles entre 2020 y 2025 relativas a la vacunación contra la COVID-19, la infección por SARS-CoV-2 y el cáncer. Concretamente, se trató de (i) categorizar las neoplasias malignas notificadas en proximidad temporal a la vacunación o la infección, (ii) evaluar los patrones temporales y clínicos en los distintos tipos de tumores para detectar señales relevantes entre los pacientes expuestos a las vacunas contra la COVID, y (iii) esbozar los mecanismos inmunológicos y moleculares plausibles que podrían subyacer a estos fenómenos.
En la bibliografía publicada, identificamos informes relacionados con neoplasias hematológicas, incluidos linfomas y leucemias, tumores sólidos como cánceres de mama, pulmón, páncreas y gliales, neoplasias asociadas a virus, como el sarcoma de Kaposi y el carcinoma de células de Merkel, y entidades poco frecuentes, como sarcomas, melanomas y carcinomas adenoides quísticos. Aunque el número de estudios o su asociación temporal no establecen una causalidad, ahora es de vital importancia comprender si estas asociaciones representan una coincidencia, una desregulación inmunitaria o un efecto biológico más amplio que vincula la infección, la vacunación y el desarrollo del cáncer.
Es importante destacar que, en lo que respecta a los efectos adversos notificados y los riesgos potenciales, el conocimiento de lo que ha ocurrido, aunque en última instancia resulte ser extremadamente raro, es un componente necesario del consentimiento informado en un momento en el que ya no existe una emergencia de salud pública por la COVID-19. Es probable que el riesgo de cáncer se base en la heterogeneidad entre las personas, el impacto de la genética, el entorno y los determinantes sociales de la salud que interactúan y varían entre las personas, y esta es un área en la que este artículo podría sentar las bases para futuros estudios que perfeccionen el riesgo individualizado. Por lo tanto, el objetivo de este artículo es sintetizar y contextualizar sistemáticamente los hallazgos de la literatura publicada sobre las neoplasias malignas asociadas temporalmente con la vacunación contra la COVID-19 o la infección por SARS-CoV-2, sin intentar estimar el riesgo, establecer la causalidad o informar las decisiones clínicas o de vacunación individuales.
Resultados
Esta revisión exploratoria, que abarca el período comprendido entre enero de 2020 y abril de 2025, no se diseñó para estimar el riesgo o la incidencia del cáncer, ni para extraer inferencias causales, sino para recopilar, categorizar y contextualizar sistemáticamente los informes publicados sobre neoplasias malignas asociadas temporalmente con la vacunación contra la COVID-19 o la infección por SARS-CoV-2. Se identificaron 69 publicaciones [8, 20–87] que describían neoplasias malignas o progresión maligna en asociación temporal con la vacunación contra la COVID-19 o la infección por SARS-CoV-2, que abarcaban un total de 333 pacientes (excluyendo los estudios a nivel de población [8, 20]). Además, se identificó una publicación a nivel poblacional que ofrecía una evaluación longitudinal de la incidencia del cáncer durante la pandemia y el período inmediatamente posterior a la pandemia [85]. Entre los 69 estudios, la mayoría de los informes eran informes de casos de un solo paciente o series pequeñas (55/69, 81 %), con un pequeño número de revisiones sistemáticas o narrativas (3/69, 4,5 %), estudios mecánicos/experimentales (2/69, 3 %) y series de casos más amplias, análisis multicéntricos o a nivel de bases de datos (8/69, 12 %) (Tabla 1 [Véase artículo original]). En consonancia con una fase temprana de detección de señales, la base empírica subyacente se inclina en gran medida hacia la documentación de casos de eventos potencialmente adversos y observaciones a nivel de casos que generan hipótesis, en lugar de estudios epidemiológicos basados en la población.
Distribución geográfica
Los informes procedían de una amplia gama de países de Asia, Europa, Oriente Medio, África y América del Norte y del Sur. Los países con mayor número de publicaciones fueron Japón (n = 11) y Estados Unidos (n = 11), seguidos de China (n = 7) e Italia (n = 4). Se identificaron casos individuales adicionales o series pequeñas en España, Corea del Sur, Arabia Saudí, India, Nigeria, Brasil, Turquía, Singapur, Líbano, Egipto, Bulgaria, Taiwán, Ucrania, Irán, Rusia, Grecia, Austria, Alemania, Polonia/Ucrania, así como colaboraciones multiinstitucionales o internacionales. Esta amplia distribución geográfica indica que las asociaciones temporales notificadas entre la vacunación o la infección por COVID-19 y los eventos oncológicos no se limitan a una región o un sistema sanitario concretos, sino que se han observado en diversos entornos clínicos e infraestructuras de diagnóstico de todo el mundo.
Tipos de exposición: vacunación frente a infección
La mayoría de las publicaciones identificadas en la búsqueda se centraron en eventos oncológicos que se produjeron después de la vacunación contra la COVID-19 (56/69; 89 %), mientras que el resto describía asociaciones tras la infección por SARS-CoV-2 (5/69; 7 %) y la infección por SARS-CoV-2 con vacunación previa (7/69; 10 %). Una publicación (1/69; 1 %) no especificaba explícitamente si el evento oncológico notificado se produjo tras la vacunación, la infección por SARS-CoV-2 o una combinación de ambas exposiciones. Entre ellas se incluían informes de casos y estudios mecánicos que evaluaban el comportamiento tumoral postinfeccioso, la perturbación inmunitaria o la aceleración de la enfermedad junto con la infección por SARS-CoV-2, pero en ausencia de vacunación o asociada a una infección por SARS-CoV-2, pero con vacunación previa o refuerzo. El predominio de los informes de casos asociados a la vacunación puede reflejar patrones de notificación más que un riesgo biológico comparativo, y los datos disponibles carecen de detalles suficientes a nivel individual para determinar si las respuestas oncológicas difieren entre la infección y la vacunación, o de qué manera lo hacen.
En toda la literatura publicada, las formulaciones de vacunas y los tipos de exposición notificados eran heterogéneos, pero podían agruparse en amplias categorías de plataformas (Figura 1). Entre los informes relacionados con las vacunas, la mayoría se referían a vacunas de ARNm, con aproximadamente el 56 % tras la vacuna de Pfizer-BioNTech (BNT162b2) y el 25 % tras la vacuna de Moderna (mRNA-1273). Otro 5 % se refería a pacientes que habían recibido ambos productos, de Pfizer y Moderna, en diferentes dosis. Las vacunas con vector adenoviral representaban la siguiente categoría más importante, incluyendo AstraZeneca (ChAdOx1/Covishield) (5,8 %), Johnson & Johnson (Ad26.COV2.S) (2,9 %) y la rusa Sputnik-V (1,4 %). Las vacunas inactivadas (por ejemplo, Sinopharm BBIBP-CorV, CoronaVac u otras formulaciones) y los estudios en los que no se informó del tipo específico de vacuna fueron los menos representados (2,6 % y 1,1 %, respectivamente). Esta distribución indica que la bibliografía publicada se inclina en gran medida hacia las plataformas de vacunas de ARNm, en particular Pfizer-BioNTech y Moderna, que juntas representan la gran mayoría de los informes relacionados con las vacunas. Este patrón refleja fielmente las prácticas de vacunación mundiales, en las que las vacunas de ARNm fueron las más utilizadas. La representación relativamente menor de las vacunas de vector adenoviral y las plataformas inactivadas probablemente refleje tanto su uso más limitado en determinadas regiones como las diferentes prácticas de notificación, más que una evaluación comparativa del riesgo biológico.
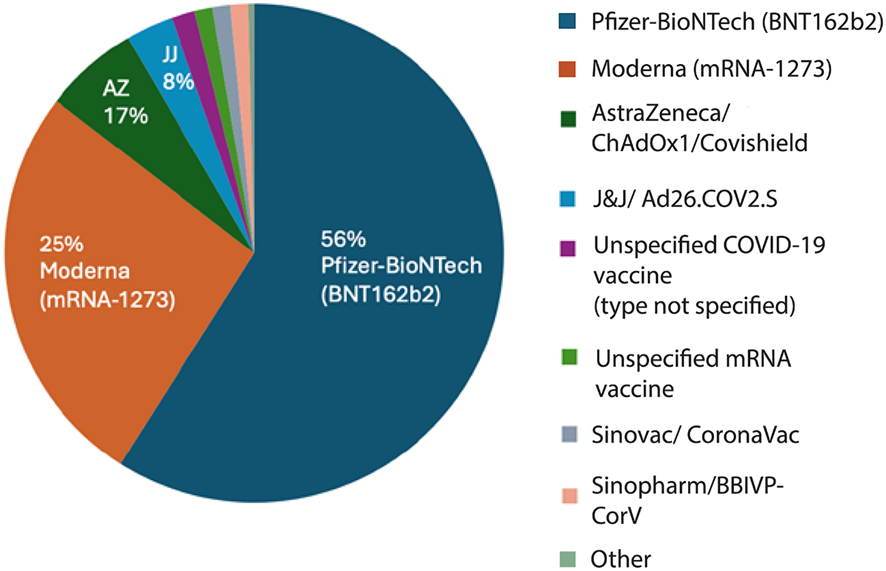
Distribución de las formulaciones de las vacunas entre los pacientes vacunados con cáncer notificado tras la inmunización contra la COVID-19. La mayoría de los casos se relacionaron con las vacunas de Pfizer-BioNTech (BNT162b2; 56 %) y Moderna (mRNA-1273; 25 %), seguidas de AstraZeneca/ChAdOx1 (Covishield; 17 %) y Johnson & Johnson/Ad26.COV2.S (8 %). Una pequeña parte de los informes se refería a Sinovac (CoronaVac), Sinopharm (BBIBP-CorV) u otras vacunas inactivadas, así como a tipos de vacunas de ARNm o COVID-19 no especificados. El predominio de las vacunas de ARNm refleja su uso generalizado a nivel mundial durante el período de estudio.
Tipos de cáncer y espectro clínico
Aproximadamente el 43 % (30/69) de las publicaciones informaron sobre neoplasias linfoides malignas, que abarcaban tanto linfomas como leucemias (Figura 2 y Tabla 2 [Véase en el artículo original]). Entre ellas se incluía un amplio espectro de neoplasias linfoides, como el linfoma difuso de células B grandes (DLBCL), diversos linfomas de células T (por ejemplo, linfoma angioinmunoblástico de células T, linfoma de células T subcutáneo similar a la paniculitis), leucemia linfocítica crónica/linfoma linfocítico pequeño (CLL/SLL) y linfomas cutáneos de células T (CTCL). Varios informes destacaron una progresión inesperadamente rápida, presentaciones atípicas o cursos inusualmente agresivos de la enfermedad.
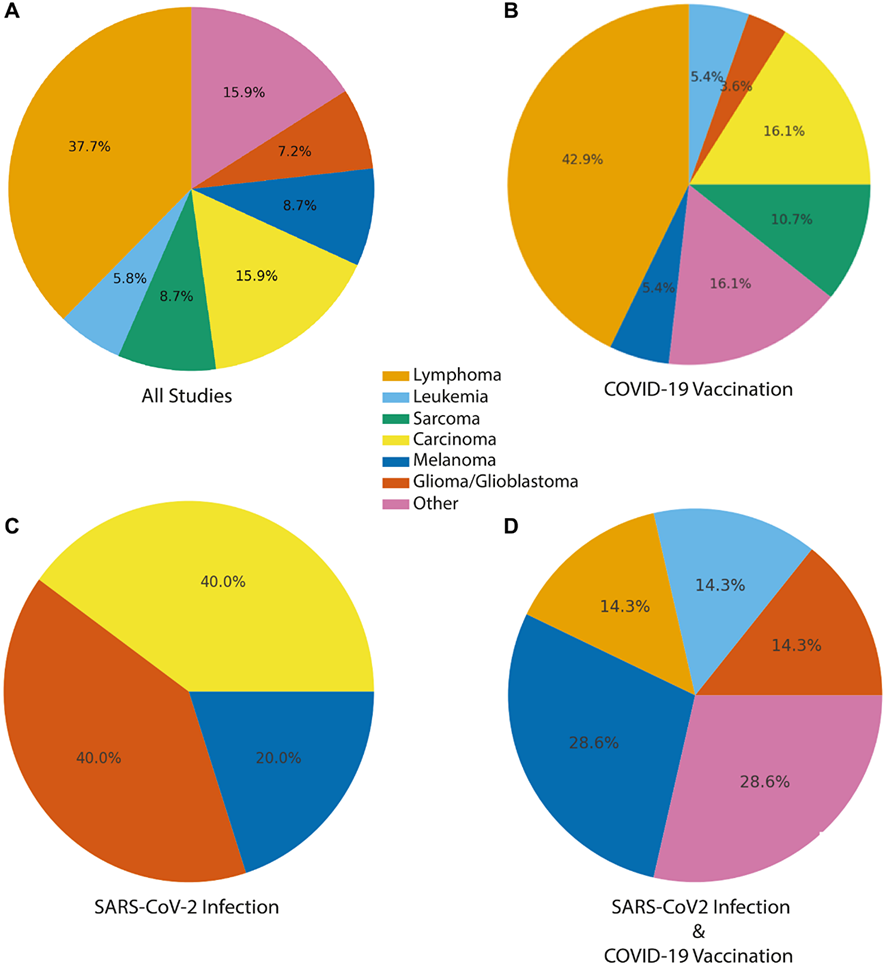
Distribución de los informes con casos de cáncer o lesiones tumorales asociados temporalmente con la vacunación contra la COVID-19, la infección por SARS-CoV-2 o la infección por SARS-CoV-2 y la vacunación. Los gráficos circulares muestran la representación proporcional de las principales categorías de cáncer observadas. (A) En todos los estudios. (B) Vacunación contra la COVID-19, (C) infección por SARS-CoV-2 y (D) infección por SARS-CoV-2 y vacunación contra la COVID-19 combinadas. Los tipos de cáncer se consolidaron en siete categorías de alto nivel. El carcinoma incluye: cáncer de mama, cáncer de próstata, cáncer de colon, cáncer de páncreas, cáncer de pulmón, carcinoma de células de Merkel, neoplasia gastrointestinal/poliposis. El linfoma también incluye neoplasias linfoides, trastornos linfoproliferativos cutáneos y trastornos linfoproliferativos. Otros incluyen tumores benignos, pseudotumores, tumores mixtos, tumores cardíacos, tumores inflamatorios y no específicos (por ejemplo, miofibroblásticos).
Los tumores sólidos representaron el 41 % de las publicaciones (28/69) y constituyeron un grupo diverso de neoplasias malignas, entre las que se incluyen el melanoma, el cáncer de mama, el cáncer de pulmón, el glioblastoma y otros tumores gliales, los sarcomas y diversos carcinomas específicos de órganos, como el cáncer de páncreas (figuras 2 y 3). En múltiples informes, los autores describieron una aparición inusualmente rápida, una recurrencia de latencia corta o una progresión clínica agresiva para tipos de tumores como el adenocarcinoma de páncreas y el glioblastoma; características atípicas para estos cánceres que se destacaron como observaciones temporales notables.
[… A continuación, en el artículo original, se describen diferentes tipos de cánceres y su asociación con la vacunación contra la Covid-19…]
Hipótesis mecánicas que relacionan la vacunación contra la COVID-19 o la infección con eventos oncogénicos
Los estudios de casos y los datos emergentes a nivel poblacional descritos anteriormente pueden representar una señal temprana de una posible asociación entre la vacunación o la infección y el cáncer que justifica una investigación más profunda. Esto plantea la pregunta: si existe una asociación, ¿cuál podría ser la base mecánica de la misma?
Los virus pueden causar cáncer [94–97]. La relación entre la infección viral y el cáncer está bien documentada en el caso del virus del papiloma humano (VPH), que causa cáncer de cuello uterino, cáncer de cabeza y cuello, así como cáncer anal, que es más frecuente entre las personas infectadas por el VIH. El virus de la hepatitis B (VHB) y el virus de la hepatitis C (VHC) causan cáncer de hígado. El virus de Epstein Barr (VEB) causa cáncer nasofaríngeo, linfoma de Burkitt y otros tipos de cáncer. El virus del herpes humano KSHV/HHV-8 causa el sarcoma de Kaposi, el virus de la leucemia de células T humanas (HTLV-1) causa leucemia o linfoma de células T en adultos, y el virus de las células de Merkle (MCV) causa cáncer de piel de células de Merkle. Se sospecha que varios virus causan cáncer, entre ellos el SV40 (mesotelioma, cánceres primarios de cerebro y huesos, entre otros) y el HCMV (glioblastoma, meduloblastoma, cáncer de mama, colon y próstata). El VIH está fuertemente asociado con el sarcoma de Kaposi, el cáncer de cuello uterino, el linfoma, el cáncer anal y otras neoplasias malignas, en gran parte debido a la inmunosupresión y la coinfección con virus oncogénicos. Se sabe desde hace décadas que las proteínas virales se dirigen a los supresores tumorales del huésped, como p53 y Rb, suprimen el sistema inmunitario y activan señales oncogénicas.
Además, las vacunas de ARNm contra la COVID actúan instruyendo a las células diana para que produzcan la proteína de espiga del SARS-CoV-2. Esto se consigue introduciendo un ARNm sintético modificado (mod-ARNm) que incorpora pseudouridina no natural en su región codificante para prolongar la estabilidad del ARNm más allá de la del ARNm natural. La introducción del mod-ARN se lleva a cabo mediante transfección lipídica en forma de nanopartículas lipídicas (LNP). El resultado es una transfección altamente eficiente del ARNm modificado en las células diana con un comportamiento bioquímico y farmacológico diferente al del ARNm natural. En consecuencia, el ARN modificado se transcribe en la proteína espiga extraña (así como en otros productos proteicos con desplazamiento del marco de lectura), lo que provoca una respuesta inmunitaria robusta [98–102]. Dada la estabilidad del ARNm modificado con pseudouridina, junto con el ADN residual en las formulaciones de las vacunas de ARNm [103–108], las vacunas de ARNm están introduciendo material genético exógeno (ADN y ARN (en forma de ácidos nucleicos modificados)) en las células de los pacientes. Las vacunas de ARNm contra la COVID-19 producen la proteína espiga, que está codificada por un ARNm estable y se ha descubierto que tiene una larga vida en el cuerpo humano [109, 110]. Se ha informado de que estos elementos de ácido nucleico contribuyen al síndrome posvacunal contra la COVID (PCVS/PVS) [110, 111]. Por lo tanto, estas vacunas se ajustan a la definición de terapia génica [112, 113]. A pesar de ello, la UE está tratando de modificar la definición de terapia génica para excluir las vacunas de ARNm de esta categoría [114].
Aunque no hay estudios que demuestren un mecanismo causal directo por el que las vacunas de ARNm induzcan cáncer, los efectos moleculares acumulativos de la proteína de pico persistente [115, 116], la activación inmunitaria y la inflamación debidas a la vacunación repetida [117–119] o la posibilidad de que se produzcan eventos de integración genómica [120] podrían contribuir a eventos que, en teoría, podrían manifestarse en forma de cánceres tras la vacunación o la infección. Dada la rápida aparición de tumores agresivos y raros que se describe en la literatura, los cánceres que surgen semanas o meses después de la vacunación serían quizás más coherentes con mecanismos que implican la promoción tumoral que con la iniciación tumoral per se. Sin embargo, también se tienen en cuenta los mecanismos que implican la iniciación. A continuación, presentamos al menos tres mecanismos biológicamente plausibles que podrían explicar la asociación entre la vacunación contra la COVID-19 y el cáncer; dos de ellos se solapan con la infección por COVID, la disfunción inmunitaria y la biología de la proteína de pico, y las reacciones debidas a impurezas del ADN restringidas a la vacunación.
Desregulación inmunitaria
La rápida aparición del cáncer, la proximidad anatómica de los tumores a los lugares de vacunación y las características histológicas de la inflamación respaldan los mecanismos inmunitarios que promueven la progresión de clones latentes en lugar de la carcinogénesis de novo. Planteamos dos procesos interrelacionados: inflamación localizada y modulación del microambiente tumoral con inmunosupresión funcional transitoria que relaja la vigilancia inmunitaria. Podría explicar la hiperprogresión de las células cancerosas latentes u ocultas (Figura 5).
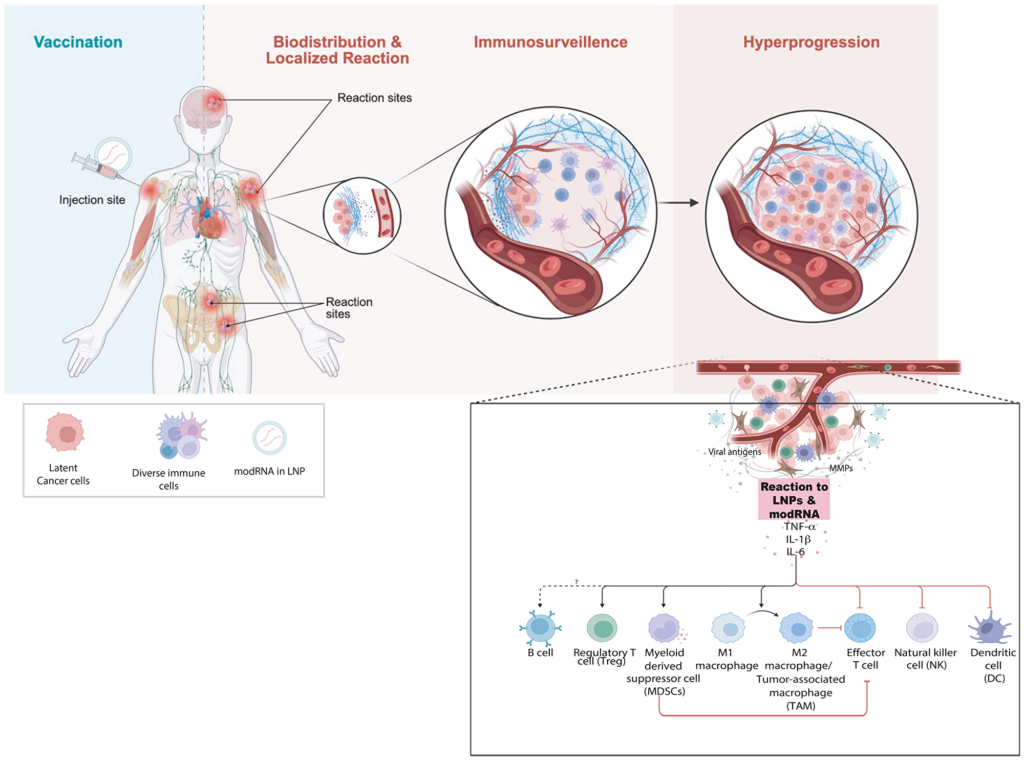
(A) Modelo conceptual que ilustra cómo la inoculación con la vacuna de ARNm provoca reacciones inmunitarias en función de su biodistribución. Una fuerte inmunoestimulación puede anular la vigilancia inmunitaria de las células cancerosas latentes y desencadenar la hiperprogresión tumoral. (B) Representación esquemática de los principales tipos de células inmunitarias que influyen en el crecimiento tumoral y la regulación inmunitaria tras la exposición a la vacuna de ARNm. El ARNm modificado encapsulado en LNP (modRNA/mRNA) interactúa con los sensores inmunitarios innatos, alterando la señalización de las citoquinas (TNF-α, IL-1β, IL-6) y la polarización de las células inmunitarias, lo que conduce a la inmunosupresión y a la reducción de la actividad de las células T CD8+ citotóxicas. La expansión de las poblaciones de supresores mieloides, junto con los bucles de retroalimentación de citoquinas pro-tumorales, fomenta la proliferación acelerada de las células tumorales y la evasión inmunitaria. El desequilibrio entre las redes antitumorales (M1, CD8+, NK) y pro-tumorales (M2, Treg, MDSC) favorece la hiperprogresión tumoral.
Numerosos estudios (tanto en humanos como en animales) han demostrado que las vacunas de ARNm contra la COVID y la infección desencadenan la producción de citoquinas proinflamatorias, entre ellas la interleucina-6 (IL-6), el TNF-α y la IL-1β, en los 1-3 días posteriores a la vacunación [121–124]. En el caso de la vacunación, la reacción se debe a la respuesta inmunitaria innata a los componentes del ARNm y las nanopartículas lipídicas (LNP), que activan los receptores de reconocimiento de patrones TLR7/8 y NLRP3 [125–128]. Por lo tanto, la producción local de estas citoquinas se producirá en cualquier lugar donde se biodistribuyan el ARNm y las LNP, lo que incluye el lugar de la inyección, los ganglios linfáticos drenantes y otros lugares distantes [129].
[…]
Más allá de la inducción innata de citoquinas, varios estudios han descrito cambios inmunitarios adaptativos transitorios tras la vacunación con ARNm o una infección aguda [110, 141–145]. Aunque estos hallazgos se interpretan generalmente como un reflejo de la inmunorregulación fisiológica y no de una disfunción inmunitaria manifiesta, pueden corresponder a períodos cortos de reducción de la respuesta inmunitaria [146–149]. Si se producen estas ventanas, ya sea de forma sistémica o localizada en nichos tisulares específicos, en teoría podrían permitir la expansión transitoria de clones virales o neoplásicos latentes. En la actualidad, sigue sin estar claro si se produce una inmunosupresión funcional transitoria después de la vacunación con ARNm y si es localizada o sistémica. Se necesitan estudios directos para determinar si la inducción de citoquinas innatas y la modulación inmunitaria adaptativa podrían contribuir o influir en los eventos cancerosos posteriores a la vacunación en individuos susceptibles.
Se han sugerido pruebas que respaldan estos mecanismos tras la propia infección por SARS-CoV-2. Por ejemplo, Gregory et al. [74] informaron de un glioblastoma agresivo con una mediana de 35 días tras la infección por COVID-19, y en un paciente se documentó que había recibido la vacuna contra la COVID-19 antes del diagnóstico. La rápida aparición del glioblastoma se atribuyó a una alteración inmunitaria que podría deberse en parte al neurotropismo del SARS-CoV-2 [150, 151] o a la biodistribución de las LNP [125, 127, 129] y a la respuesta inmunitaria a estas. De hecho, la proteína espiga se ha localizado en el tejido cerebral, así como en las células de glioblastoma y los macrófagos que rodean las células tumorales [87]. Hu et al. [72] demostraron de manera similar que la exposición viral directa y la presencia de la proteína espiga, con lesión mediada por citoquinas en organoides de glioma, aumentaban la proliferación y la invasividad de las células tumorales, lo que respalda un modelo de estimulación tumoral impulsada por la infección. En el caso del cáncer de mama, Chia et al. [74] demostraron que las infecciones virales respiratorias, incluido el SARS-CoV-2, podían despertar rápidamente las células cancerosas metastásicas latentes en el pulmón a través de la activación impulsada por el interferón del eje STAT1-NF-κB, que remodela el nicho local a un estado pro-metastásico. Tanto el virus como las vacunas de ARNm activan los sensores inmunitarios innatos y provocan respuestas complejas de citoquinas e interferones que pueden remodelar la interfaz tumor-inmunitaria.
En conjunto, estas observaciones sugieren que la activación inflamatoria aguda, la refractariedad inmunitaria de corta duración y los lapsos transitorios en la vigilancia citotóxica forman un marco biológicamente plausible a través del cual la vacunación podría influir y promover el comportamiento de las células neoplásicas preexistentes o latentes. El mecanismo inmunológico sugiere que la infección y la vacunación pueden operar a lo largo de un continuo biológico común, que difiere principalmente en la intensidad, la biodistribución y la persistencia de las perturbaciones inmunitarias y moleculares que inducen. Aunque aún no se dispone de pruebas causales directas, la convergencia de los aumentos innatos de citoquinas, la modulación transitoria de la dinámica de las células T y B y las señales asociadas a la regulación inmunitaria ponen de relieve la importancia de seguir investigando. Aclarar la magnitud, la duración y la especificidad tisular de estos estados inmunitarios posteriores a la vacunación será esencial para determinar si, y en qué individuos, tienen relevancia clínica para la progresión o la recurrencia del cáncer.
Biología de la proteína espiga
La transformación de una célula normal en una célula cancerosa implica la alteración de múltiples mecanismos de protección que controlan el crecimiento celular, la supervivencia y la reparación del ADN. Estudios de laboratorio han demostrado que la proteína espiga, ya sea producida por infección o por vacunación, tiene actividades biológicas [110, 145, 152–158] con potencial oncogénico [159–161]. Por ejemplo, además de interactuar con los receptores ACE2, se ha demostrado que los fragmentos de la proteína espiga interactúan con NRP-1, integrinas y TLR, lo que conduce a la señalización VEGF/NRP-1 [155, 162, 163]. También se ha informado de que la proteína espiga induce daños en el ADN [160, 164, 165] y modifica la vía p53 bajo estrés oxidativo [164, 166]. Por lo tanto, en teoría, estas interacciones de la proteína espiga con estas vías podrían contribuir a la transformación celular, tanto de la vacuna como de la infección, especialmente si la proteína espiga permanece presente mucho tiempo después de la vacunación o de múltiples infecciones por COVID.
Se ha informado de que la proteína espiga producida por las vacunas persiste durante semanas, meses e incluso años [109, 110, 116, 167–170] después de la vacunación, lo que proporciona una actividad potencial a largo plazo en las células. Además, la proteína espiga estabilizada producida por las vacunas contra la COVID (Spike-2P) difiere de la proteína natural del SARS-CoV-2, ya que contiene dos sustituciones de prolina (K986P y V987P) que permiten su estabilización [171]. Por ello, será importante evaluar si la incidencia del cáncer se correlaciona con la expresión (o persistencia) de la proteína espiga variante en el organismo, pero también si es esta versión la que está presente en los tumores, como se ha descrito en gliomas y astrocitomas [73], así como en cáncer de mama metastásico [36]. En un caso reciente de cáncer de mama metastásico en la piel, la lesión apareció un mes después de la sexta dosis de la vacuna (Pfizer-BioNTech) y las células cancerosas metastásicas se tiñeron con la proteína espiga, pero no con la proteína nucleocápside del virus SARS-Cov-2 ([36], Figura 3F), lo que sugiere que se trataba de la proteína espiga derivada de la vacuna. Por lo tanto, la exposición crónica a un agente con actividad biológica que altera el ciclo celular y las vías de respuesta al daño del ADN podría representar un nuevo factor etiológico del cáncer. Cabe destacar y es relevante para el glioblastoma u otras patologías del sistema nervioso central (SNC) tras la infección por COVID o la vacunación el tropismo de la proteína espiga hacia el SNC [150, 151, 172].
Contaminantes del ADN
El ADN residual en los productos biológicos es un subproducto bien establecido y reconocido de la fabricación de vacunas, con límites establecidos por la FDA y la Organización Mundial de la Salud (OMS), pero solo para el ADN desnudo, no para el ADN encapsulado en LNP [173]. Las impurezas de ADN en las vacunas de ARNm surgen debido al subproducto de la transcripción in vitro [174] y pueden incluir ADN de doble cadena (ADNdc), ARN abortivos e híbridos de ARN:ADN [103, 174]. Están encapsulados por nanolípidos, lo que permite un acceso más estable y eficiente a las células, lo que aumenta el riesgo de integración [128, 175, 176]. Además, el ADN residual en las formulaciones de vacunas de ARNm [103–108] procedente del proceso de fabricación supera los límites establecidos incluso para el ADN desnudo. Algunos estudios han comparado directamente la eficiencia de transfección del ADN desnudo con la del ADN encapsulado en LNP y han demostrado que la integración de la transfección basada en lípidos es significativamente mayor que la del ADN desnudo [175]. Además, es bien sabido que los músculos esqueléticos y cardíacos absorben (e incluso expresan ADN plasmídico desnudo) in vivo [177–179]. En particular, un estudio de tumores cardíacos en el período posterior a la COVID reveló un aumento del 1,5 % en la incidencia de tumores y la expresión de la proteína de espiga en los tumores, especialmente en los asociados a la vacunación [86].
La cantidad de ADN residual comunicada en varias evaluaciones independientes supera los límites reconocidos para el ADN desnudo, y la distribución del tamaño de los fragmentos de ADN, cuando se combina con una mayor eficacia de transfección debido a las LNP, plantea la posibilidad de una inserción genómica. Además, dado que los elementos reguladores del SV40 están presentes en las impurezas de la vacuna BNT2b [180], cuando se inserta en el genoma, este ADN puede alterar la expresión de secuencias adyacentes y/o la regulación normal de los genes y aumentar el potencial tumorigénico [120, 181]. El ADN extraño, especialmente cuando se administra en LNP altamente inflamatorias [182], puede activar vías de detección inmunitaria innatas, como el cGAS-STING citosólico y el receptor Toll-like 9 (TLR9) endosomal, lo que da lugar a respuestas de interferón tipo I y citoquinas inflamatorias [183, 184].
Los límites de impurezas del ADN se establecieron para el ADN desnudo [185], no para el ADN encapsulado en LNP, que provoca una mayor absorción celular y persistencia intracelular de los fragmentos de ADN. Esto aumentará la posibilidad de mutagénesis por inserción, lo que puede provocar reordenamientos genómicos, así como la integración y expresión de proteínas de pico persistentes, la alteración de la regulación génica normal y la posible activación de vías protooncogénicas o la inactivación de supresores tumorales. De hecho, los estudios in vitro demuestran tasas de integración genómica de entre el 1 % y el 10 % de las células transfectadas inicialmente con sistemas de administración basados en lípidos [120]. No se han realizado estudios que demuestren que el nivel de impurezas de ADN presentes en las vacunas sea insuficiente para transfectar las células, ni se han realizado estudios que descarten la posibilidad de integración.
Lagunas en el conocimiento
A pesar de la escala global sin precedentes de la vacunación contra la COVID-19, siguen existiendo profundas lagunas en nuestra comprensión de cómo las plataformas de vacunas de ARNm interactúan con las vías fundamentales de defensa del huésped, la homeostasis tisular y la biología tumoral. Estas lagunas abarcan la biología a nivel molecular, celular, orgánico y poblacional. No hay datos que relacionen las vacunas de ARNm, y en especial las vacunas de ARNm contra la COVID-19, con las consecuencias biológicas posteriores.
A nivel molecular, existen importantes lagunas de conocimiento sobre cómo las modificaciones químicas y estructurales de la proteína de espiga del SARS-CoV-2, las sustituciones de nucleósidos y aminoácidos (por ejemplo, N1-metilpseudouridina) y las formulaciones de LNP influyen en la señalización de las células huéspedes, la estabilidad genómica y la regulación inmunitaria. La ingeniería de vacunas se ha centrado en maximizar la inmunogenicidad antiviral, pero se sabe mucho menos sobre las posibles interacciones colaterales entre la expresión de la proteína espiga, la supresión tumoral, el daño al ADN o las vías de respuesta al estrés que podrían modularse inadvertidamente durante una activación inmunitaria intensa o una señalización celular alterada que afecte a las defensas del huésped contra el cáncer.
La distinción entre la iniciación y la promoción de tumores asociadas a la vacunación también sigue sin resolverse. No existe una validación empírica de que la vacunación solo acelere la enfermedad preexistente en lugar de iniciar también nuevas neoplasias. Dado que las mutaciones somáticas y las células neoplásicas latentes son omnipresentes en los tejidos adultos, la aparición de tumores de latencia corta en un plazo de semanas a meses puede reflejar la promoción de clones latentes en lugar de una carcinogénesis de novo, un fenómeno coherente con la hiperprogresión observada en subgrupos de pacientes que reciben inhibidores de puntos de control inmunitario [138, 139]. Los factores oncogénicos, como la amplificación de MDM2/MDM4 o la amplificación, sobreexpresión o mutaciones de EGFR, se han relacionado con la hiperprogresión [138, 140, 186] y la agresividad metastásica [187, 188], lo que sugiere que los cambios en las citoquinas o los puntos de control inducidos por las vacunas podrían, en teoría, converger en vías oncogénicas similares. Tampoco está claro cómo las vacunas de ARNm dirigidas contra el SARS-CoV-2 podrían estar sensibilizando los tumores a los inhibidores de puntos de control inmunitario, como se ha sugerido recientemente [19].
[…]
Conclusiones
Las pruebas colectivas a nivel mundial de 2020-2025 subrayan una conexión biológicamente plausible entre la vacunación contra la COVID-19 y el cáncer. Los hallazgos clínicos recurrentes documentados en muchos informes sobre la aparición de cáncer de novo, la rápida progresión tumoral, la reactivación viral y el despertar de enfermedades latentes ponen de relieve lagunas críticas en el conocimiento y la comprensión de cómo los cambios inmunitarios a gran escala producidos por la vacuna interactúan con la biología del cáncer.
Tanto la infección por SARS-CoV-2 como la vacunación contra la COVID-19 activan vías biológicas superpuestas que, en principio, podrían influir en el riesgo de cáncer, pero difieren en cuanto al mecanismo, la magnitud, la biodistribución y la duración de sus efectos. Los mecanismos comunes incluyen la activación del sistema inmunitario innato, la señalización robusta del interferón, la inducción de citoquinas, el estrés oxidativo y la alteración transitoria de la homeostasis de las células inmunitarias. En teoría, estos cambios pueden exponer neoplasias malignas latentes, promover la expansión clonal de células mutantes preexistentes o crear contextos microambientales que favorezcan la progresión tumoral.
Además, tanto la infección como la vacunación inducen la expresión de la proteína espiga, que interactúa con los tejidos que expresan ACE2 y puede desencadenar la activación endotelial, la inflamación y las vías de estrés celular implicadas en la señalización oncogénica. Ambas pueden también conducir a estados prolongados de inflamación y lesión tisular, lo que podría contribuir a la inestabilidad genómica, la remodelación epigenética y la desregulación inmunitaria crónica.
Sin embargo, hay mecanismos únicos que distinguen la vacunación contra la COVID-19 de la infección natural. La vacunación implica una biodistribución generalizada, la captación intracelular y la persistencia de plantillas de ácido nucleico modificadas que impulsan la síntesis de una proteína espiga no natural tanto en el lugar de la inyección como en todo el organismo. La presencia de ADN residual o fragmentado, combinada con la administración mediada por LNP a tejidos inmunitarios y no inmunitarios, y la expresión sostenida de la proteína Spike durante meses o años, representan factores específicos de la vacuna que, en teoría, podrían promover la mutagénesis insercional, perturbar la vigilancia inmunitaria o acelerar el crecimiento de clones malignos preexistentes. Por lo tanto, hay mucho que aprender de las muestras de tejido y sangre humanos, así como de las autopsias, para comprender mejor la interacción entre la infección por COVID, la vacunación y los mecanismos del cáncer.
Con este fin, es necesario estudiar la presencia y persistencia de la proteína espiga, junto con los efectos biológicos que son autónomos de las células o que dependen de las interacciones inmunitarias del huésped, para establecer conexiones con la iniciación y progresión del cáncer. En consecuencia, proponemos que los tumores que surjan después de una infección documentada por SARS-CoV-2 o tras la vacunación contra la COVID-19 se evalúen utilizando un marco de clasificación inmunohistoquímica estandarizado.
Como mínimo, esto debería incluir la evaluación de los patrones de expresión de antígenos virales mediante IHC. Deben definirse los fenotipos positivo para la proteína de espiga/positivo para la nucleocápside, positivo para la proteína de espiga/negativo para la nucleocápside y negativo para la proteína de espiga/negativo para la nucleocápside. Esta evaluación debe integrarse con una caracterización detallada de la actividad proliferativa (por ejemplo, Ki-67), los marcadores de respuesta a la muerte celular y al daño del ADN, las firmas de las vías supresoras de tumores y oncogenes, y el microambiente inmunitario del tumor.
La implementación de este tipo de informes en las evaluaciones de patología clínica y autopsias permitiría una discriminación más precisa entre los tumores potencialmente impulsados por la infección, por la expresión de antígenos relacionados con la vacuna o por procesos oncogénicos no relacionados, y permitiría la agregación de casos comparables entre instituciones. El establecimiento de criterios uniformes de este tipo es esencial para construir una base de evidencia coherente, apoyar la investigación mecanicista y, en última instancia, determinar si las asociaciones observadas reflejan una coincidencia, el desenmascaramiento de una enfermedad latente, una perturbación inmunitaria o verdaderas relaciones causales.
Establecer la causalidad entre la infección por SARS-CoV-2, la vacunación contra la COVID-19 y el cáncer requiere un nivel de evidencia mucho más allá de la asociación temporal. En oncología, la causalidad nunca se determina mediante una sola observación o estudio, sino que surge solo cuando múltiples líneas de evidencia independientes convergen a lo largo del tiempo. Esto incluye datos mecanicistas (como análisis de integración genómica, trayectorias de evolución clonal, perfiles inmunológicos y cambios epigenéticos), hallazgos basados en la patología (incluidas autopsias con caracterización molecular), modelos experimentales que reflejan con precisión la biología de los tejidos humanos (organoides, sistemas humanizados, secuenciación de lectura larga de tejidos expuestos) y estudios epidemiológicos a nivel de población capaces de detectar señales pequeñas pero significativas frente a la incidencia de fondo. Solo integrando estos enfoques podemos distinguir la coincidencia del desenmascaramiento de una enfermedad latente, la expansión de clones malignos preexistentes o la verdadera oncogénesis de novo. Es importante destacar que la necesidad de pruebas rigurosas no debe utilizarse para descartar patrones emergentes.
Es esencial un debate transparente sobre los mecanismos biológicamente plausibles y las estrategias de vigilancia para determinar si esta asociación temporal tiene una relación causal. Se carece de datos epidemiológicos fiables actuales que proporcionen pruebas de que la vacunación no aumenta la incidencia de cáncer a nivel poblacional. La literatura revisada por pares no está indexada de forma completa ni fácil. El establecimiento de un marco para la vigilancia del cáncer tras la vacunación podría ayudar a detectar precozmente patrones adversos poco frecuentes y permitir un seguimiento mecánico sin comprometer la confianza del público. El objetivo de esta revisión no es estimar el riesgo de cáncer a nivel poblacional, sino proporcionar una síntesis estructurada de la literatura revisada por pares existente, identificar temas clínicos y biológicos recurrentes y delinear las lagunas críticas que requieren un seguimiento epidemiológico y mecánico riguroso. Esto permitirá comprender mejor todo el espectro de respuestas inmunitarias para informar estrategias de inmunización más seguras y arrojar luz sobre los vínculos entre la inmunidad y la biología del cáncer que antes se subestimaban.
El imperativo científico para avanzar debe ser un marco coordinado que combine la vigilancia longitudinal y la experimentación mecánica para permitirnos distinguir la coincidencia de la causalidad y perfeccionar las futuras plataformas de vacunas en consecuencia. Al hacerlo, no solo obtendremos una comprensión más clara de la seguridad de las vacunas, sino también una visión más profunda de los vínculos fundamentales entre la inmunidad, la infección y la aparición del cáncer.
CONTRIBUCIONES DE LOS AUTORES
C.K. y W.E-D. concibieron la revisión, realizaron la búsqueda bibliográfica, la extracción de datos y la síntesis, redactaron y aprobaron la versión final del manuscrito.
AGRADECIMIENTOS
W.S.E-D. es profesor investigador de la Sociedad Americana contra el Cáncer y cuenta con el apoyo de la Cátedra Universitaria de la Familia Mencoff en la Universidad de Brown.
CK es director del Laboratorio de Convergencia de Ciencias Biomédicas, Físicas y de Ingeniería de la Universidad de Tufts.
CONFLICTOS DE INTERÉS
W.S.E-D., coeditor jefe de Oncotarget, no participó en la revisión de este manuscrito ni en la decisión de aceptarlo.
NOTA EDITORIAL
El editor jefe, el Dr. Wafik S. El-Deiry, no participó en el proceso de revisión por pares ni en la toma de decisiones sobre este artículo. El Dr. El-Deiry compartió el manuscrito enviado con el director del Instituto Nacional del Cáncer (NCI), Anthony Letai, por correo electrónico el 12 de diciembre de 2025.
Referencias: https://www.oncotarget.com/article/28824/text/
————————-
Un resumen de este estudio se puede encontrar en: https://cancerintegral.com/vacuna-covid-arma-bioterrorista-primer-estudio-revisado-por-pares-que-la-relaciona-con-el-incremento-de-cancer/
————————–