Por el Dr. Jonathan Latham, y la Dra. Allison Wilson, 17 de marzo de 2024
Este texto se publicó originalmente como un hilo en Twitter/X.
Lo volvemos a publicar (salvo algunos errores tipográficos) como un breve ensayo.
1/ Mucha gente en Twitter se pregunta, como Nicholas Wade, si las últimas informaciones sobre la subvención DEFUSE de @emilyakopp y @USRTK son quizás la última palabra en el gran debate sobre el origen del #covid #lableak. No lo son.
2/ ¿Por qué? Porque no hacen más que ahondar en el misterio. La respuesta corta es que seis sitios distintivos de ingeniería inversa identificados por Bruttel et al: (https://www.biorxiv.org/content/10.1101/2022.10.18.512756v1) sólo explican el 0,1% o menos del genoma del #SARS-CoV-2.
3/ Si aceptamos provisionalmente que demuestran un #lableak, y la evidencia indica que deberíamos hacerlo, ¿de dónde, sin embargo, procede el resto del genoma? ¿No sería interesante saberlo?
4/ Podría serlo, porque aunque el SARS-CoV-2 muestra pruebas de manipulación, TAMBIÉN muestra pruebas igualmente claras de evolución y selección para un huésped humano. ¡Una contradicción contundente!
5/ La evidencia más reciente de esa evolución se puede encontrar en un artículo de julio de ’23 (Ou et al, 2023) que obtuvo pocas lecturas, casi ninguna cita, y prácticamente ningún interés en Twitter por lo que podemos decir. Lo encontramos la semana pasada. Probablemente, fue ignorado en parte porque es de China.
6/ Ahora bien, China seguramente tiene censores científicos, por lo que es necesario darle credibilidad, pero la mayoría de los experimentos de Ou et al pueden corroborarse con pruebas generadas fuera de China. Aquí hay un enlace al artículo de Ou et al: https://www.nature.com/articles/s41421-023-00581-9
7/ Evadió la censura probablemente porque en China suponen que este trabajo prueba un origen evolutivo y refuta así un #lableak. Se trata de un error lógico que, no obstante, debemos agradecer. Aquí están las conclusiones. Es un gran trabajo con múltiples conclusiones clave.
8/ (Por cierto, los investigadores proceden de prestigiosas instituciones de Pekín, el inglés es excelente y los razonamientos y la lógica son en su mayoría cristalinos). Así que no se lo pierdan. ADEMÁS, enumeramos las conclusiones por orden de magnitud creciente, así que intente leer hasta el final.
9/ La idea básica de la investigación es observar a los parientes más cercanos del SARS-CoV-2 (Específicamente BANAL-52 y BANAL-236, encontrados por investigadores franceses/laosianos en el norte de Laos) y compararlos en cuanto a estructura y función, y a veces incluyendo al SARS1.
10/ Se centraron específicamente en 1) la unión del receptor (ACE2) a humanos y murciélagos; 2) las secuencias de aminoácidos (AA); 3) las estructuras tridimensionales de las proteínas; y 4) la eficacia de entrada del virus en las células. (En adelante, SARS-CoV y SARS-CoV-2 se denominarán aquí SARS1 y SARS2).
11/ Hay 3 resultados principales:
RESULTADO 1: Unión de SARS1, SARS2, BANAL-52 y BANAL-236 (en adelante B-52 y B-236) a animales huéspedes putativos.
12/ Los PRINCIPALES resultados: La unión de la espiga SARS2 fue más intensa en humanos. La unión del SARS2 a la ACE2 del perro mapache fue bastante débil. Después de los humanos, la unión más fuerte de SARS2 fue a las ACE2 de pangolín y gato de algalia (ver Fig. 1.).
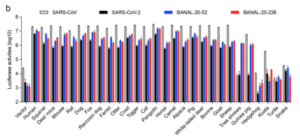
13/ Obsérvese también que, con la unión ACE2 del perro mapache, tanto el SARS1 como el B-52 y el B-236 se unen mejor que el SARS2, que se queda unas 5 veces por detrás (nótese la escala logarítmica). En todo caso, un escenario de zoonosis del perro mapache esperaría que el SARS2 se uniera MEJOR, no menos, que el ancestro.
14/ Los experimentos anteriores necesitan ser reproducidos de forma independiente (¿pero por quién?), pero sería dar una mala imagen para la tesis favorita actual de la zoonosis (y puede explicar por qué ningún laboratorio occidental ha informado de este obvio, clave y fácil, para así tener que decir «Nunca lo sabremos»).
15/ RESULTADO 2 Los autores descubrieron que las espigas B-52 y B-236 se unían bien (o muy bien) a las 14 ACE2 de murciélagos rinolófidos analizadas. Por el contrario, SARS2 se unía menos fuertemente que B-52 y B-236 a todas las ACE2 de murciélagos y muy débilmente a algunas. SARS2 SIEMPRE se unió menos que B-52 o B-236 (véase la Fig. 2).
16/ Esta débil unión de SARS2 a bat ACE2s se debe principalmente a una única diferencia AA: H498Q . SARS2 tiene una Q (Glutamina) en la posición 498 mientras que las BANAL llevan una H (Histidina).
17/Pruebas de ello:
Las espigas enteras de B-52 y SARS2 difieren en sólo 19 aminoácidos (AAs) y sólo en esa única posición 498 en el dominio RBM (la parte que se une a ACE2).
18/ Los autores comprobaron el efecto de esta sustitución. Cuando se cambió SARS2 por H se unió MUCHO MEJOR a algunas ACE2 de murciélago (no todas parecen haber sido probadas). Por el contrario, el cambio de H498 a Q en B-52 redujo MUCHO la unión a las mismas ACE2 de murciélago (véase la Fig. 3).
19/ Potencialmente, este cambio fue una simple casualidad que se produjo en un cambio de huésped, ya que el cambio NO afecta a la unión con la ACE2 humana y, por lo tanto, no fue producto de la selección por ese motivo. Pero hay más.
20/ Los autores también generaron anticuerpos contra cada proteína de la espiga. Luego probaron los Abs generados contra B-52 para determinar su actividad contra el SARS2 CON Y SIN el cambio Q498H. Tener una Q hizo que el SARS2 fuera 2,2 veces menos sensible a los Abs (ver Fig. 4). Esto es mucho para una sola diferencia de AA.
21/Su conclusión fue que el 498H es altamente inmunogénico. Estiman que la Q del SARS2 le permite evadir el 25% de los Abs contra el B-52. Puede que sólo sea casualidad. Sin embargo, este hallazgo ofrece una razón por la que el SARS2 perdió su H (que se conserva en todos los RBM de coronavirus de murciélagos estrechamente relacionados, es decir, los de pangolines y BANAL-103) en un nuevo huésped.
22/ PERO tal explicación sólo es relevante en huéspedes con una fuerte inmunidad adquirida (como pacientes de larga duración o poblaciones inmunes/vacunadas). No hay muchas teorías de contagio que encajen. Pero véase más abajo.
23/RESULTADO 3: Aquí se llega al meollo de la cuestión: Los autores descifraron la estructura tridimensional de los picos B-52 y B-236. En la conformación cerrada, ambos eran muy similares. En la conformación cerrada, ambos eran muy compactos en comparación con el SARS2.
24/ Ambas tenían también un glicano extra (carbohidratos unidos a las proteínas, a menudo para protección física) en la posición 370. Esta pérdida de glicano ya era conocida: https://pubs.rsc.org/en/content/articlehtml/2021/sc/d1sc04832e
25/ El SARS2 carece de este glicano debido a una diferencia de Treonina a Alanina en la posición 372 (T372A) de la espiga del SARS2. (Esta T forma el tercer AA en un motivo (NXT) necesario para la glicosilación, por lo que el glicano se une a la N en la posición 370 aunque la mutación esté en la 372).
26/ Esta glicosilación también está muy conservada entre los betacoronavirus. Incluso el MERS y el SARS1 la tienen, al igual que todos sus parientes cercanos. Probablemente es importante para ellos (Ver Fig. 5.).

27/ Anteriormente, Kang et al habían demostrado que la eliminación de este glicano de la espiga del SARS2 aumenta su afinidad por el receptor humano ACE2 en 20 veces. Eso sí que es importante. Kang: https://www.sciencedirect.com/science/article/pii/S0092867421008333
28/ ¿Por qué todos estos virus murciélago renuncian a ese grado de mejora de la unión al receptor? ¿Qué hace el T372 por los virus murciélago para que merezca la pena conservarlo?
29/ Recordemos que los coronavirus de murciélago son TODOS patógenos del tracto gastrointestinal y que los picos B-52 y B-236 eran compactos. Resulta que la simulación del intestino delgado de los murciélagos (con pH 5,5 y añadiendo la proteasa tripsina) hace que los mutantes A372 de B-52 y B-372 se desintegren.
30/ En resumen, el glicano 370 protege la espiga, que está en la superficie del virus, de la degradación en el intestino del murciélago, pero esta protección física es a costa de reducir la unión del receptor de la espiga. El mecanismo específico es probablemente que la posición abierta de la espiga (la posición «arriba»), que se requiere para la unión del receptor, también la expone a la digestión.
31/ Así pues, la elección de T o A en la posición 372 representa un compromiso, y en el pulmón, donde las proteasas son mucho menos abundantes y el pH es más neutro, A se ve favorecido sobre T. Así pues, el glicano 370 no es necesario en los pulmones y el virus puede aprovechar una conformación más abierta.
32/ Así pues, el SARS2 está adaptado a los pulmones. Lo he comprobado, y hacen falta dos cambios de nucleótidos para que la T ancestral en cualquiera de los ancestros conocidos se convierta en una A en el SARS2. Por lo tanto, tienen que producirse dos mutaciones sucesivas una al lado de la otra. Eso supone un considerable grado de evolución.
33/ De las dos mutaciones, una es sinónima (no cambia el AA) y la otra es no sinónima (cambia el AA). En una población de virus, las mutaciones no sinónimas pueden cambiar la frecuencia muy rápidamente, pero las mutaciones sinónimas suelen cambiar la frecuencia sólo lentamente.
34/ Ou et al ofrecen pistas muy limitadas sobre lo que piensan; Pero aquí está lo que pensamos:
35) 1) El resultado del perro mapache (RESULTADO 1) es intrigante como un argumento más razonable pero no concluyente contra una zoonosis por perros mapaches.
36) 2) La clara implicación del RESULTADO 3* es que la mutación del Glicano 370 no ocurrió en murciélagos (donde sería altamente deletérea). Podría haber mutado en un hospedador intermediario, pero habría llevado un tiempo considerable.
37/ Un tiempo prolongado en una zoonosis implica una cadena de infección sustancial pero inadvertida en huéspedes intermedios. Ocurrió con el SRAS1 pero dejó evidencias.
38/ Muchos animales infectados deberían haberse visto, ya sea 1) por su enfermedad, O, 2) a través de múltiples contagios. Recordemos que el SARS1 se propagó en animales y pasó a humanos múltiples veces. Lo mismo ocurrió con el MERS. Sin embargo, en este caso sólo hay pruebas de UN ÚNICO contagio.
39/ Recordemos también que la espiga del SARS2 PERDIÓ afinidad por la espiga del perro mapache en comparación con sus ancestros BANAL. Si éstos eran el huésped intermediario y, sin embargo, la espiga pasó un largo período de tiempo en estos animales, ¿por qué perdería gran parte de su afinidad por su ACE2?
40/ El RESULTADO 3, sin embargo, es consistente con nuestra teoría del Mojiang Miner. Para las personas no familiarizadas se describe aquí https://www.independentsciencenews.org/commentaries/a-proposed-origin-for-sars-cov-2-and-the-covid-19-pandemic/
41/ En pocas palabras, propusimos que el SARS2 evolucionó en uno de los seis mineros que contrajeron una misteriosa enfermedad similar al COVID en 2012. Teorizamos que, al igual que omicron, el coronavirus de murciélago que probablemente tenían, evolucionó dentro de ellos dadas sus hospitalizaciones prolongadas.
42/ El WIV (lo sabemos) recogió múltiples muestras de ellos. El WIV ha negado desde entonces que estas muestras contuvieran un coronavirus, pero esto es discutible: https://www.independentsciencenews.org/commentaries/a-chinese-phd-thesis-sheds-important-new-light-on-the-origin-of-the-covid-19-coronavirus/
43/ Lo que nos resulta especialmente interesante es que la evolución de un virus de murciélago a SARS2 dentro de un único huésped humano es TOTALMENTE COMPATIBLE CON LOS RESULTADOS 1, 2 Y 3. La teoría del minero ofrece lo que una #zoonosis probablemente no puede: evolución fuera de un murciélago y en presencia de inmunidad adquirida.
44/ Las implicaciones para un escenario de #lableak son un poco diferentes. La ingeniería genética por sí sola no creó el SARS2. No se trató de un simple caso de ingeniería inversa de un esqueleto de murciélago o incluso de añadirle un sitio de furina. Se requieren demasiados pasos.
45/ Si fueran necesarias múltiples mutaciones además de un sitio furina para que el SARS2 infectara los pulmones humanos tan bien como lo hace, ¿quizás el posterior pasaje y selección en cultivos celulares o modelos animales suplieran la diferencia?
46/ Existen precedentes del paso de coronavirus manipulados https://link.springer.com/article/10.1186/1743-422X-8-64
Sin embargo, son considerablemente menos sofisticados de lo que se requeriría en este caso. Parece que se necesitaría competencia inmunitaria, humanización ACE2, pulmones y un largo período de tiempo.
47/ Predecimos, por tanto, que 2024 podría ser un buen año para la teoría de la Evolución del Minero de Mojiang (#MMPtheory). Es la manera más fácil (parsimoniosa) de conciliar todos estos datos, además de todas las pruebas que ya existen, y tal vez la única manera.
48/ En primer lugar, voy a presentar un póster sobre ello en la próxima Conferencia de la Sociedad de Virología en Viena, a partir del 25 de marzo. Estaré en el puesto 429 el lunes 25 de marzo. #GfV2024
49/ Debería ser interesante, sobre todo teniendo en cuenta el tumulto de la última vez que fui a una conferencia de virología:
*Nota: Yuri Deigin nos señaló, (a través de Twitter) que la solicitud de subvención DEFUSE tiene una discusión sobre la alteración de glicanos en los picos de los coronavirus murciélago que encuentran. El pico del SARS2 tiene 22 glicanos. La discusión de la subvención DEFUSE es algo vaga; no menciona el glicano en N370 específicamente, ni tampoco los documentos que citan, y las alteraciones propuestas fueron por otras razones, ya que la importancia del glicano 370 es nueva información, pero su discusión acerca de los glicanos plantea la posibilidad de que esto también podría haber sido manipulado.
—————–